Залози та їх гормони в організмі
У людському організмі крім основних функціональних органів знаходяться так звані залози, тобто органи покликані синтезувати і виділяти активні речовини, які називаються секретами. Найчастіше залози мікроскопічні і лише деякі з них мають відносно великі розміри. Що цікаво, залози, які мають вивідні протоки і виділяють секрет на слизову оболонку або поверхню тіла (потові, слинні, молочні та інші) називають екзокринними, а не мають проток і виділяють високоактивні речовини (гормони) безпосередньо в систему кровообігу (щитовидна залоза, надниркові залози , гіпофіз та інші) отримали назву ендокринними. Слід докладніше розглянути залози і їх гормони.
Що знаходиться попереду гортані на шиї людини виробляє багаті йодом гормони - тироксин, трийодтиронін і кальцитонін. Дані гормони покликані регулювати вуглеводний, жировий, а також білковий обмін в організмі, та до того ж впливають на зростання людини і його психічний розвиток. При гіперфункції даної залози у людини розвивається ожиріння, підвищується температура тіла і знижується обмін речовин.
Являють собою невеликі освіти над нирками, що складаються з двох шарів зовнішнього (коркового), а також внутрішнього (мозкового). Зовнішній шар виробляє статеві гормони, які відповідають за розвиток статевих ознак і регулюють обмін речовин (білки і жири, вуглеводи, калій, натрій). При дисфункції кори надниркових залоз людина захворює, так званої бронзової хворобою, тобто тіло його набуває бронзовий колір, а сам він відчуває постійну втому, втрату апетиту і нудоту. У функції мозкового шару надниркових залоз входить вироблення таких гормонів, як норадреналін і адреналін. Так, адреналін посилює частоту серцевих скорочень і прискорює кругообіг крові, мобілізуючи всі сили організму в стресовій ситуації. Нестачі у цього гормону не буває, а ось його надлишок загрожує звуженням кровоносних судин і прискореним серцебиттям. Навпаки ж, норадреналін покликаний сповільнювати частоту скорочень серця.
Що знаходиться в черевній порожнині підшлункова залоза є залозу змішаної секреції, так як має вивідні протоки і виділяє секрет в шлунок, беручи участь в травленні. Один з вироблюваних нею гормонів, глікоген синтезує глюкозу, підвищуючи тим самим рівень цукру в крові. Інший же гормон - інсулін, навпаки, є єдиним, який знижує рівень цукру в крові. Саме через порушення функціонування підшлункової залози з'являється цукровий діабет, при якому відбувається підвищення цукру в крові.
Статеві залози, тобто жіночі яєчники і чоловічі насінники, також є залозами змішаної секреції. За рахунок їх зовнішнього секрету з'являються статеві клітини, тобто сперматозоїди і яйцеклітини, а їх ендокринна функція полягає у виробленні чоловічих, а також жіночих статевих гормонів. Дані гормони впливають на статеву поведінку і фігуру людини певної статі. Чоловічі насінники виробляють андрогени, які відповідають за наявність чоловічих статевих ознак, мускулатури, вусів, бороди, а жіночі яєчники виробляють естрогени, що відповідають за статеві ознаки представниць прекрасної статі, тобто розвиток молочних залоз і форми тіла.
Розташовується в головному мозку гіпофіз складається з 3 частин (передній, проміжної і задньої), дві з яких виробляють гормони. В передній долі виробляється гормон, що впливає на зростання кісток і прискорює процеси обміну речовин. Його недолік може загрожувати розвитком карликовості, а гіперфункція може привести до гігантизму. Проміжна частка гіпофіза виробляє гормон, який відповідає за пігментацію шкіри.
Все вищеописане лише підтверджує, що залози і їх гормони вкрай важливі для людського організму і без них людина не змогла б існувати. Біжіть себе!
Популярне на сайті
Інформація на нашому сайті носить пізнавальний і освітній характер. Однак дана інформація ні в якій мірі не є посібником з самолікування. Обов'язково проконсультуйтеся з лікарем.
Щоб зрозуміти, як функціонують ендокринні органи, і зокрема щитовидна залоза, необхідно коротко розглянути механізм дії гормонів.
Мал. 1. Схема розташування ендокринних органів
Ендокринну функцію організму забезпечують системи, в які входять:
1) ендокринні залози, секретирующие гормони;
2) гормони і різні шляхи їх транспортування;
3) відповідні органи або тканини-мішені, що відповідають на дію гормонів.
Ендокринна система підтримує сталість внутрішнього середовища організму, необхідне для нормального перебігу фізіологічних процесів.
Залози внутрішньої секреції є спеціалізовані органи, які мають залозисте будова. Розрізняють залози тільки з внутрішньою секрецією (гіпофіз, надниркові залози, щитовидна залоза, околощітовідние залози) і змішані - з внутрішньої і зовнішньої секрецією. Прикладом може служити підшлункова залоза. Її зовнішня секреція полягає у виробленні травних ферментів, які за спеціальним протоку надходять в дванадцятипалу кишку, а внутрішня секреція полягає в тому, що в спеціалізованих бета-клітинах панкреатичних острівців (Лангерганса) виробляється гормон інсулін, що надходить безпосередньо в кров і регулює рівень цукру в крові . Статеві залози також здійснюють внутрішню і зовнішню секрецію.
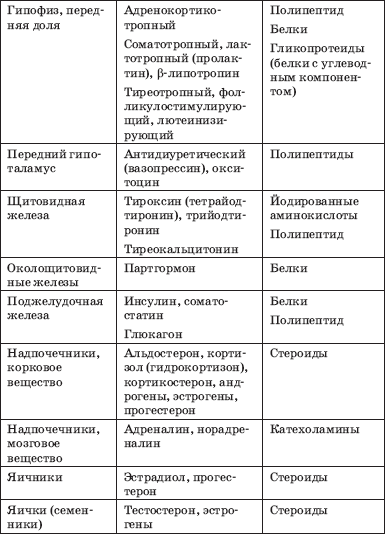
Назва і розташування залоз внутрішньої секреції, які продукують ними гормони, хімічна природа останніх представлені в табл. 1.
Таблиця 1. Гормони залоз внутрішньої секреції (Потьомкін В. В., 1986)

Закінчення табл. 1
Термін «гормон», в перекладі з грецького означає «я буджу», «спонукаю», був введений в практику Бейлісс і Старлингом. У січні 1902 вони провели свій знаменитий, що став класичним досвід, який переконливо довів участь гуморального фактора в регуляції секреторної діяльності підшлункової залози. Бейлісс і Старлінг вважали гормоном будь-яка речовина, в нормі продукується клітинами будь-якої частини організму і переноситься кров'ю до віддалених частинах, на які воно діє для блага організму в цілому.
В даний час гормони визначають як високоактивні речовини, що утворюються в залозах внутрішньої секреції, що надходять в кров і надають регулюючий вплив на функції віддалених від місця їх секреції органів і систем організму. Їх називають також хімічними посередниками, які секретуються безпосередньо в кровотік спеціалізованими клітинами, здатними синтезувати і вивільняти гормони у відповідь на специфічні сигнали.
За хімічною будовою гормони діляться на:
1) гормони - похідні амінокислот;
2) білкові і поліпептидні гормони;
3) прийом препарату.
За фізіологічною дією гормони ділять на пускові та виконавці. До пусковим гормонів (активаторам діяльності інших ендокринних залоз) відносяться нейрогормони гіпоталамуса і тропний гормони гіпофіза. Гормони-виконавці надають безпосередній вплив на основні функції організму.
Гормони відрізняються від інших біологічно активних речовин наступними властивостями:
1) дуже високою біологічною активністю;
2) дистанційним характером дії;
3) суворої специфічністю.
Висока біологічна активність гормонів характеризується тим, що, перебуваючи в крові в мізерно малих кількостях, вони мають виражену дію.
Дистанційний характер дії гормонів полягає в тому, що точки прикладання їх дії розташовуються зазвичай далеко від місця утворення гормону в ендокринній залозі.
Гормони відрізняються строгою специфічністю дії. Це означає, що реакції органів, тканин і клітин на гормони строго вибагливі. Кожен гормон впливає тільки в певних органах і тканинах, так званих органах-мішенях (тканинах-мішенях). Гормон дізнається і взаємодіє зі своїм органом-мішенню тому, що в цих органах є спеціальні з'єднання - рецептори. Рецептори - це інформаційні білкові молекули, що розпізнають і трансформують гормональний сигнал в гормональне дію. До теперішнього часу ідентифіковано більше 60 рецепторів. Для стероїдів (гормонів кори надниркових залоз) і тиреоїдних гормонів (гормонів щитовидної залози), легко проникають через мембрану, рецепторні білки розташовані всередині клітини. Рецептори для білкових гормонів і катехоламінів, які не можуть пройти через клітинну мембрану, розташовані на поверхні клітини.
Гіпоталамус і гіпофіз є єдиною системою управління периферійними ендокринними залозами.
Гіпоталамус - це частина мозку, що володіє властивостями нервової і ендокринної систем. В гіпоталамус надходить великий потік інформації від органів чуття і внутрішніх органів. До складу нейросекреторну ядер гіпоталамуса входять так звані крупноклеточние і дрібноклітинні ядра. Перші виділяють гормони окситоцин і вазопресин, які по нервових стовбурах транспортуються в задню частку гіпофіза, накопичуються там і в міру потреби використовуються для регуляції діяльності нирок і матки.
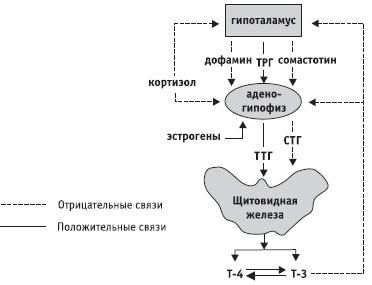
Мал. 2. Схема регуляції гіпоталамо-гіпофізарно-тиреоїдної системи
Інші функції виконують дрібноклітинні ядра гіпоталамуса. Вони здатні виробляти так звані рилізинг-гормони, або, правильніше, рилізинг-фактори (дозволяють фактори). Рилізинг-фактори по венозній системі досягають гіпофіза і регулюють виділення гормонів останнього.
Регуляція діяльності гіпофіза гормонами дрібноклітинних ядер гіпоталамуса здійснюється по антагоністичному принципом. Одна група чинників стимулює виділення гормонів гіпофіза (рилізинг-фактори, або ліберіни), а інша - гальмує (статини). Відомі такі чинники: кортиколиберин, стимулюючий секрецію адренокортикотропного гормону гіпофіза; тіроліберін, що підсилює виділення тиреотропного гормону гіпофіза; соматолиберин і соматостатин (перший стимулює виділення соматотропного гормону гіпофіза - гормону росту, а другий - гальмує); меланоліберін і меланоцітостатін і ін.
Гіпофіз є центральною ендокринної залозою, в якій виробляються так звані гормони тропів, що регулюють функцію периферичних залоз. Це складний ендокринний орган, розташований в підставі головного мозку - в так званому турецькому сідлі. Він складається з аденогіпофіза, більшу частину якого становить передня частка залози, і нейрогипофиза, представленого його задньої часткою.
В передній долі (аденогипофизе) виробляються гормони тропів:
Гормон росту, який регулює процеси росту організму, синтезу білка, глюкози і розпаду жиру;
Кортикотропін, стимулюючий синтез глюкокортикоїдів в корі надниркових залоз;
Тиреотропін - стимулятор синтезу тиреоїдних гормонів щитовидної залози;
Гонадотропін, фоллікулотропін, що регулюють синтез чоловічих і жіночих статевих гормонів;
Пролактин - гормон, що регулює лактацію.
У задній частині гіпофізу (нейрогіпофіз) скупчуються вазопресин і окситоцин. Вазопресин, або антидіуретичний гормон, регулює водний обмін і тонус судин. Окситоцин підвищує тонус гладкої мускулатури матки, регулює родовий акт і виділення молока грудними залозами.
Периферичні ендокринні залози діляться на дві групи.
Першу складають залози, функція яких регулюється тропними гормонами аденогіпофіза. Їх називають аденогіпофіззавісімимі залозами, або залозами-мішенями. До них відносяться щитовидна залоза, кіркова речовина надниркових залоз, ендокринні частини статевих залоз. Взаємини між аденогипофизом і залозами-мішенями грунтуються на принципі «зворотного зв'язку». Наприклад, аденогипофиз виділяє в кров тиреотропний гормон, який стимулює виділення гормону щитовидної залози - тироксину. Поступив в кров тироксин пригноблює виділення тиреотропного гормону гіпофіза.
Другу групу ендокринних периферичних залоз складають залози, функція яких не залежить від діяльності гіпофіза. Ці залози називають аденогіпофізнезавісімимі. Вони функціонують в автономному режимі. До них відносяться околощітовідние залози, ендокринна частина підшлункової залози, мозкову речовину надниркових залоз, ендокринні клітини вилочкової залози.
В тимусі (вилочкової, або зобної, залозі) виробляються гормони тимозин і тимопоетин - стимулятори імунних процесів.
Щитовидна залоза продукує йодовмісні гормони: тироксин і трийодтиронін, а також тиреокальцитонин. Тироксин і трийодтиронін регулюють основний обмін, тобто той рівень енерговитрат, який необхідний для підтримки життєдіяльності організму в стані повного спокою. Тіреокальціто-нин регулює обмін кальцію і фосфору.
У околощитовідних залозах виробляється паратгормон, який також регулює кальцієвий і фосфорний обмін. Але якщо тиреокальцитонин щитовидної залози знижує вміст кальцію в крові, то паратгормон околощітовідних залоз підвищує його. Антагоністичні взаємини між Тиреокальцитонін і паратгормоном забезпечують вміст кальцію в крові на потрібному для організму рівні.
Виключно важлива роль гормонів наднирників. Це парні органи, розташовані над верхніми полюсами нирок. У наднирниках розрізняють кіркова і мозкова речовина.
Корковаречовина виділяє групу стероїдних гормонів, що іменується збірною назвою кортикостероїди. Три зони коркового речовини спеціалізовані на виділенні різних гормонів. Клітини клубочкової зони продукують мінералокортикоїди: дезоксикортикостерон і альдостерон, що регулюють мінеральний обмін. Пучкова зона виробляє глюкокортикоїди: кортизол і кортикостерон, що здійснюють регуляцію обміну білків, жирів і вуглеводів. У сітчастій зоні синтезуються деякі попередники чоловічих статевих гормонів (андрогенів).
Мозкова речовина надниркових залоз виділяє в кров катехоламіни - адреналін і норадреналін. Норадреналін виступає в ролі не тільки гормону, але і медіатора нервових процесів симпатичного відділу нервової системи. Катехоламіни мають виражену судинозвужувальну дію, підвищуючи тим самим артеріальний тиск. Вони беруть участь в регуляції вуглеводного і жирового обміну, відіграють основну роль в адаптації організму під час стресу. Адреналін виділяється у відповідь на найрізноманітніші подразники: страх, хвилювання, біль, радість. Його образно називають аварійним гормоном, гормоном емоцій, першим медіатором стресу.
У ендокринної частини підшлункової залози (острівцях Лангерганса) виробляються інсулін, глюкагон, соматостатин. Інсулін є найважливішим регулятором вуглеводного, а також жирового і білкового обміну. Глюкагон - це фізіологічний антагоніст інсуліну, а також стимулятор його секреції в присутності глюкози. Соматостатін пригнічує секрецію інсуліну, глюкагону і гормону росту. Порушення секреції інсуліну і глюкагону веде до розвитку такого важкого і поширеного захворювання, як цукровий діабет.
Статеві залози продукують не лише гормони, але також і статеві клітини (сперматозоїди і яйцеклітини). У сім'яниках (яєчках) виробляються чоловічі статеві гормони - андрогени, головним з яких є тестостерон. Андрогени сприяють розвитку первинних і вторинних чоловічих статевих ознак. В яєчниках синтезуються жіночі статеві гормони - естрогени, що відповідають за формування жіночих первинних і вторинних статевих ознак, а також прогестерон - гормон, необхідний для нормального перебігу вагітності. Вироблення гормонів і статевих клітин здійснюється під контролем гонадотропних гормонів аденогіпофіза.
Нирки, виконуючи функцію виділення, є також і своєрідною ендокринної залозою. Клітини так званого юкстагломерулярного апарату нирок секретують в кров гормон ренін, який бере участь в утворенні ангіотензину II - найактивнішого регулятора тонусу судин. У нирках виробляється також еритропоетин - гормон, що стимулює утворення еритроцитів в кістковому мозку.
Встановлено, що і серце є ендокринної залозою. В передсерді синтезується натрійуретичний гормон, що впливає на виділення натрію нирками.
Тимчасово функціонуючим ендокринних органом є плацента ( «дитяче місце»). У ній виробляються гормони, що сприяють нормаль ному протіканню вагітності.
У центральній нервовій системі утворюються особливі речовини - нейроен докрінние пептиди (нейрогормони) - ендорфіни, енкефаліни. Їх називають «ендогенними опіатами» або морфіноподібними пептидами. Ці гормони надають аналгезирующее (знеболююче) дію і відтворюють поведінкові ефекти морфіну.
Єдність і взаємозв'язок нервових і ендокринних механізмів дуже чітко простежуються на прикладі функціонування гіпоталамо-гіпофізарної системи. В даний час правильніше говорити не про ендокринної, а про нейроендокринної системі організму.
Виклавши загальні уявлення про залозах внутрішньої секреції, перейдемо до головної мети нашої розповіді - щитовидній залозі.
Питання №2 Ендокринні залозиі їх гормони.
ендокринні залози(Від грец. Endon - всередині, crio - виділяю) або залози внутрішньої секреції, є спеціалізовані органи або групи клітин, основна функція яких полягає у виробленні та виділення у внутрішнє середовище організму специфічних біологічно активних речовин. Залози внутрішньої секреції не мають вивідних проток. Їх клітини обплетені рясної мережею кровоносних і лімфатичних судин, і продукти життєдіяльності виділяються безпосередньо в кров, лімфу, тканинну рідину. Ця особливість принципово відрізняє ендокринні залози від екзокринних, які виділяють свої секрети через вивідні протоки.
Продукти, що виробляються залозами внутрішньої секреції, отримали назву гормонів(Грец. Hormao - збуджую, активують). Термін «внутрішня секреція» був запропонований в 1885 році французьким фізіологом К. Бернаром, а термін «гормон» - англійськими фізіологами У. Бейлісом і Е. Старлінг в 1902 році.
Для гормонів характерні наступні особливості: Синтез і виділення їх здійснюється спеціалізованими клітинами. Гормони утворюються в залізистих ендокринних клітинах, після чого вони надходять у внутрішнє середовище, в основному в кров і лімфу. Проміжні продукти синтезу або метаболізму гормонів нерідко мають біологічну активність, але, як правило, не серкетіруются .
Висока біологічна активність.Гормони роблять фізіологічна дія в дуже малих концентраціях. Так, концентрація жіночого статевого гормону (естрадіолу) в крові коливається від 0,2 до 0,6 мкг (Ю -6 г) в 100 мл плазми. Зміст гормону росту в крові вимірюється ще меншими величинами - наніграммамі (КГ 9 г). Гіпофіз реагує на пікограм (Ю -12 г) гіпоталамічних гормонів, ангіотензин-П-продукт ендокринних клітин нирок - викликає відчуття спраги в фемтограмм (Ю -15 г). Крім гормонів, ніякі інші хімічні продукти життєдіяльності не ефективні в таких малих дозах.
Специфічність.Кожен гормон характеризується певними, притаманними тільки йому хімічною структурою, місцем синтезу і функцією. У зв'язку з цим дефіцит будь-якого гормону не може бути заповнений іншими гормонами або біологічно активними речовинами.
Дистантность дії.Гормони, як правило, переносяться кров'ю далеко від місця освіти, впливаючи на віддалені органи і тканини. Цим вони відрізняються від медіаторів і цитокінів, що діють на одну клітку або групу клітин на місці їх утворення.
Хімічна структура гормонів і їхперетворення в організмі
За хімічною будовою гормони можна розділити на 4 основні групи: білки і пептиди, похідні амінокислот, стероїди, простагландини.
Прикладами білкових гормонів: інсулін, соматотропін (гормон росту), гормони тропів передньої долі гіпофіза. Деякі з них (фоллітропін, тиреотропин, лютропін) відносяться до складних білок, інші (інсулін, кальцитонін та ін.) - до простих білків. Пептидную структуру мають глюкагон, вазопресин, окситоцин, гіпоталамічні гормони. До похідних амінокислот відносяться гормони щитовидної залози - трийодтиронін, тироксин, а також адреналін і норадреналін. Стероїдні гормони мають в своїй основі ядро \u200b\u200bциклічного вуглеводню циклопентанпергідрофенантрену. До цієї групи належать гормони кори надниркових і статевих залоз.
Основні етапи освіти і перетворення гормону можна представити таким чином:
біосинтез гормону;
секреція, тобто виділення з ендокринної клітини;
транспорт кров'ю до периферичних тканин;
розпізнавання гормонального сигналу клітинами-мішенями;
трансдукция (переклад) гормонального сигналу в біологічний відповідь;
гасіння гормонального сигналу.
Органи і тканини з инкреторной функцією клітин
|
|
|
|
Органи і тканини
|
|
|
|
|
|
плацента
|
синцитіотрофобласти цитотрофобласт
|
Хоріонічний гонадотропін, пролактин Естріол, прогестерон
|
|
|
|
|
тимоцити
|
Тимозин, тимопоетин
|
|
|
|
|
інтерстицій канальці
|
еритропоетин кальцитриол
|
|
|
|
|
міоцити передсердь
|
Атріопептід, соматостатин, ангіотензин-П
|
|
|
|
|
|
|
|
Органи, тканини і клітини з ендокринною функцією
|
|
|
|
структури
|
Тканини, клітини
|
гормони
|
|
|
|
гіпоталамус
|
Нервові клітини мелкоточечних структур
|
Гіпоталамічні нейрогормони:
|
|
|
|
|
заднього гіпоталамуса Нервові клітини крупноклітинних ядер переднього гіпоталамуса
|
а) ліберіни: кортиколиберин, тіроліберін, люліберін, фолліберін, соматолиберин, пролактоліберін, меланоліберін; б) статини: пролактостатін, меланостатин, соматостатин Вазопресин, окситоцин
|
|
|
|
аденогіпофіз нейрогіпофіз
|
Кортікотрофи гонадотрофов тіреотрофов соматотрофов лактотрофи Пітуіціти
|
Кортикотропін, меланотро- пін Фоллітропін, лютропін Тиреотропін Соматотропин Пролактин Вазопресин, окситоцин
|
|
|
|
Наднирники Корковаречовина мозкова речовина
|
Клубочковая зона Пучкова зона Сітчаста зона Хромафині клітини
|
Мінералокортикоїди Глюкокортикоїди Статеві стероїди: андрогени, естрогени Андреналін, норадреналін
|
|
|
|
Щитовидна залоза
|
фолікулярні тиреоцитах
|
Трийодтиронин, тетрайодтіронін Кальцитонин, катакальцін, Ко-кальцігенін
|
|
|
|
околощітовідние залози
|
Головні клітини К-клітини
|
паратирин Кальцитонин
|
|
|
|
|
Пінеоціти
|
мелатонін
|
|
|
|
Підшлункова залоза
|
Острівці Лангерганса: ос-клітини Р-клітини А-клітини
|
глюкагон соматостатін
|
|
|
|
Статеві залози (насінники, яєчники)
|
Клітини Лейдіга клітини Сертолі клітини гранульози жовте тіло
|
Тестостерон Естроген, ингибин Естрадіол, естрон, прогестерон прогестерон
|
|
|
|
|
|
|
|
біосинтез гормонівзапрограмований в генетичному апараті спеціалізованих ендокринних клітин. Отже, він залежить від структури і експресії генів, що кодують синтез цих гормонів, а також від ферментів, що регулюють синтез гормону і Посттрансляційні процеси. Відсутність або дефект відповідних генів призводить до ендокринопатії. Прикладом може служити карликовість при генетичному дефекті гормону росту.
2.Секреція гормонів.найважливіша ознака будь-якого гормону - його секретируемой. Упакований в везикули або гранули гормон транспортується у напрямку до цитоплазматичної мембрани. Виходу білково-пептидних гормонів і катехоламінів з клітки передує взаємодія цитоплазматичної мембрани і мембрани секреторною гранули. Після цього відбувається їх лізис і вихід гормону з клітки. Даний процес активується багатьма факторами медіаторами, високою концентрацією калію, електричними стимулами і ін. Секреція гормону - акт, супроводжуваний витратою енергії, тому вона завжди пов'язана зі зрушеннями в системі АТФ - цАМФ. Для секреції необхідна участь іонів кальцію, які активують білки мікротубулярної-мікрофіламентною системи, сприяючи взаємодії з ними гранул гормону, і впливають на утворення цАМФ. Тому зниження вмісту іонів кальцію в позаклітинній середовищі і надходження його в ендокринну клітку неминуче призводить до зменшення секреторної активності цієї клітини.
3.
Транспорт гормонів.Секретується гормон потрапляє у внутрішнє середовище організму, переважно в кров, і транспортується далі. Більшість гормонів утворює в крові комплексні сполуки з білками плазми. Частина цих білків - специфічні транспортні протеїни (наприклад, транскортином, що зв'язує гормони кори надниркових залоз), частина - неспецифічні (наприклад, у-глобуліни). Комплексирование з білками - оборотний процес. Крім того, частина гормонів пов'язана в крові з форменими елементами, зокрема еритроцитами.
Освіта пов'язаної форми гормонів має велике фізіологічне значення. По-перше, це оберігає організм від надмірного накопичення в крові (і, отже, впливу на тканини) вільних гормонів. По-друге, пов'язана форма гормону є його фізіологічним резервом. По-третє, зв'язування з білками сприяє захисту гормону від руйнування ферментами, тобто продовжує його життя. Нарешті, комплексування з білками перешкоджає фільтрації мелкомолекулярних гормонів через ниркові клубочки і тим самим утримує ці важливі регуляторні процеси.
4.
Розпізнавання гормонального сигналу.Після надходження в периферичні органи гормони, як правило, звільняються від білкового компонента і, фіксуючи на певних рецепторах клітин, що сприймають цей гормон (клітинах-мішенях), здійснюють своє специфічне дію. В процесі периферичної дії гормонів відбуваються їх різні перетворення. При цьому можливе утворення нових гормональних продуктів, нерідко більш активних або виробляють інше біологічне дію, ніж вихідний гормон. Так, гормон щитовидної залози тироксин може в тканинах перетворюватися в трийодтиронін - більш активний гормон цієї ж залози. Андрогени (чоловічі статеві гормони) перетворюються в гіпоталамусі в естрогени - жіночі статеві гормони. Трансдукція гормонального сигналу в біологічний відповідь органічно пов'язана з механізмом дії цього гормону.
Дія будь-якого гормону на клітини-мішені завжди починається з взаємодії його з певними компонентами клітини. Це явище називається рецепцією гормону, а клітинні компоненти, які взаємодіють з гормоном, - рецепторами. Рецепторами гормонів є кислі крупномолекулярні олігопептиди.
Структура рецепторной молекули характеризується асиметричністю. Виділяють три її ділянки:
зв'язує гормон;
еффекторний, передає гормональний сигнал на внутрішньоклітинні механізми, тобто здійснює трансдукцію сигналу в біологічний відповідь;
сполучають перший і другий ділянки.
- гіпофіз синтезує / секретує соматотропний гормон (СТГ), пролактин, АКТГ і ін .;
- в наднирнику - чотири шари клітин, кожен з яких синтезує свій гормон.
Підшлункова залоза, з точки зору гастроентеролога, - екзокринний орган, так як вона секретує панкреатичні ферменти; з точки зору ендокринолога, - це ендокринний орган, так як вона продукує пакет взаємозалежних гормонів (інсулін, глюкагон, соматостатин та ін.).
Крім того, деякі гормони утворюються в декількох місцях:
- катехоламіни - не тільки в мозковому шарі надниркових залоз, а й в паравертебральних нервових гангліях;
- соматостатин - і в острівцях Лангерганса і в гіпоталамусі.
Поза ендокринних залоз виявлені мікроскопічні скупчення клітин, що спеціалізуються на синтезі біологічно активних речовин з властивостями гормону:
- регулятори секреції гормонів ендокринних залоз:
- ядра гіпоталамуса синтезують речовини, що регулюють секрецію гормонів гіпофіза (соматолиберин, АКТГ-рилізинг-гормон і ін.);
- скупчення клітин в стінці кишечника, які продукують гормони інкретіна;
- регулятори функцій органів:
Відносно недавно виявлені біологічно активні речовини лептин і адипонектин, синтезовані жировою тканиною (адипоцитами), які були віднесені до гормонів, так як володіють системною регулюючим дією - регулюють апетит і енергообмін.
Отже, гормони продукує не тільки ендокринна заліза, внаслідок чого це якість не може однозначно визначати поняття «гормон». Разом з тим в сучасній клінічної ендокринології практично всі хвороби є те чи інше порушення функцій саме ендокринної залози. У зв'язку з цим визначення гормону і пов'язане з ним визначення ендокринної залози в клінічній ендокринології все ще залишається «класичним».
Таким чином, можна дати наступне, досить повне з клінічної точки зору визначення гормону.
гормон - біологічно активна речовина, що виробляється ендокринної залозою, яке надає регулюючу дію на певні структури організму і обмін речовин (утилізацію субстратів з крові, енергообмін і ін.), Що часто проявляється зовні видимими змінами організму (наприклад, зростання) і / або зміною поведінки ( наприклад, статевий).
У цьому класичному визначенні терміни ендокринна заліза і гормон взаємозалежні. Звідси очевидна в клінічній ендокринології логіка діагностичного пошуку - через дослідження гормонів крові діагностувати хвороби ендокринних залоз.
Визначення ендокринної залози
ендокринна залоза - чітко окреслена макроанатоміческая структура, головною функцією якої є синтез біологічно активних речовин, які називаються гормонами. У клінічній ендокринології виділяють сім ендокринних залоз, функції яких оцінюють по дослідженню в крові виробляються залозою гормонів. Для оцінки її функцій використовують не весь спектр гормонів залози, а строго обмежений їх набір, за допомогою якого визначають функцію ендокринної залози. Крім гормонів, для діагностики хвороб можна використовувати їх метаболіти, які іноді виявляються більш надійним маркером ендокринної хвороби, ніж дослідження самих гормонів. Так, в діагностиці феохромоцитоми надійніше дослідження метаболітів катехоламінів метанефринів, ніж адреналіну і норадреналіну.
Дослідження гормонів для діагностики ендокринних захворювань не завжди виправдано. Найбільш яскравий приклад - цукровий діабет, в діагностиці якого не використовують дослідження інсуліну, хоча захворювання викликане інсуліновою недостатністю. Також дослідження окситоцину і вазопресину не використовують для діагностики їх недостатньої або надмірної секреції, а порушення їх синтезу визначають по їх метаболічним ефектам.
Більш того, в діагностиці ендокринних хвороб можуть бути використані гормони, які не синтезуються ендокринними залозами, наприклад інсуліноподібний ростовий фактор I (ІРФ-I), який утворюється в печінці під дією СТГ. Його використовують для діагностики акромегалії, викликаної пухлиною гіпофіза.
Синтез гормону ендокринної залозою може бути:
- єдиною її функцією (наприклад, передня частка гіпофіза);
- суміщений з генеруванням статевих клітин (наприклад, яєчники і тестікули);
- суміщений з екзокринної секрецією (наприклад, підшлункова залоза);
- суміщений з депонуванням гормонів, що синтезуються за її межами.
Ендокринна залоза здатна синтезувати:
- єдиний гормон, що зустрічається рідко (наприклад, паращитовидної залози);
- спектр гормонів (як правило):
- спеціалізованими клітинними Субструктура, зокрема в надниркових залозах дві клітинні субструктури - кора і мозковий шар - виробляють стероїдні гормони і катехоламіни відповідно;
- окремими клітинами, об'єднаними чи ні в ізольовані комплекси, наприклад в гіпофізі певні гормони синтезують окремі клітини, що не об'єднані в виразні клітинні освіти; в підшлунковій залозі інсулін і глюкагон продукуються β- і α-клітинами, об'єднаними в острівці Лангерганса.
Природа і функції гормонів
Гормони поділяють на дві основні групи.
Поліпептиди або похідні амінокислот (більшість):
- складні поліпептиди (ЛГ, ХГЧ);
- пептиди середніх розмірів;
- малі пептиди;
- дипептиди (Т 4 і Т 3);
- похідні окремих амінокислот (серотонін, гістамін).
Похідні холестерину - стероїди двох типів:
- з інтактним стероїдних кільцем (стероїди наднирників і гонад);
- з роз'єднаним кільцем В.
Виділяють чотири основні функції гормонів в організмі:
- розмноження;
- зростання і розвиток;
- виробництво, утилізація та збереження енергії.
Окремий гормон, з одного боку, може надавати різні біологічні ефекти на різні органи, а в одному органі-в різний час; з іншого боку, деякі біологічні процеси перебувають під інтегральним контролем декількох гормонів.
Гормони регулюють функції наступних своїх мішеней:
- інші ендокринні залози (наприклад, зв'язок гіпофіз-надпочечник);
- функціональні системи;
- органи (наприклад, Т 4 і функція серця або Т 4 і функція мозку);
- тканини (наприклад, кортизол і кісткова тканина).
Синтез, зберігання і секреція гормонів
Пептидні гормони синтезуються за тим же механізмом, що і будь-які інші білки. Часто спочатку синтезується велика молекула прогормона, яка потім перетворюється в гормон менших розмірів. Наприклад, препропаратіреоідний гормон → пропаратіреоідний гормон → паратиреоїдного гормон. З іншого боку, стероїди і катехоламіни синтезуються з менших молекул.
Ендокринні органи не є унікальним місцем синтезу гормонів, разом з тим лише в них синтез гормонів і його регулювання відбуваються найбільш ефективно. Три основні особливості відрізняють ендокринний орган від неендокріннимі тканини, що синтезує будь-якої гормон:
- швидкість синтезу набагато вище в ендокринній органі;
- ендокринні залози забезпечені механізмом транспорту гормону в кров, який, як правило, регулюємо.
Швидкість секреції гормону залозою визначається швидкістю його синтезу, яку можуть регулювати інші, тропів по відношенню до цієї залозі гормони. За винятком Т 4 і 1,25-дігідроксіхолекальціферол, запаси гормонів в організмі дуже обмежені.
Стимуляція секреції гормону пов'язана з деполяризацією клітинної мембрани і відкриттям кальцієвих каналів, що призводить до вступу кальцію в клітину, де він з'єднується з кальцій-зв'язуючим протеїном.
Транспорт і елімінація гормонів
Гормони елімінуються з крові в результаті метаболічних процесів, наприклад пептидні гормони інактивуються протеолітичнимиферментами. У печінки гормони з'єднуються з глюкуроновою кислотою і секретуються з жовчю, але при цьому частково реабсорбуються, включаючись в так званий ентеропечёночний цикл. Гормони також виводяться з сечею.
Невеликі молекули гормонів (Т 4, зокрема) зв'язуються з білками крові, що уповільнює їх елімінацію з крові і підтримує невеликий пул вільного гормону в крові на необхідному рівні. Зв'язування з білками також полегшує транспорт жиророзчинних стероїдів.
гормональні рецептори
Рецептори гормонів - клітинні білки, які зв'язують гормон.
Взаємодія з гормоном викликає в рецепторі конформационное зміна, що активує специфічну клітинну ферментну систему, яка фактично і реалізує характерний ефект гормону. Коли гормон з'єднується з рецептором клітинної мембрани, в цитоплазмі з'являються так звані другі месенджери (перший - гормон). У ядрі клітини гормон-рецепторний комплекс пов'язаний з дезоксирибонуклеїнової кислотою (ДНК) і регулює експресію гена. Максимальний ефект гормону зазвичай проявляється вже в тому випадку, якщо пов'язано менше 50% рецепторів. Вивільнені від зв'язку з гормоном вільні рецептори знову повертаються в цитозоль або на мембрану клітини, де продовжують брати участь в гормон-рецепторном взаємодії.
Стероїдні гормони ліпофільних, тому вільно дифундують через клітинну мембрану, а потім зв'язуються з цитозольними рецепторними білками.
Т3 зв'язується з ядерними рецепторними білками, і комплекс Т 3-рецептор, з'єднуючись з ДНК, стимулює утворення матричної РНК. Нерідко стероїдні і тиреоїдні гормони діють синергічно, взаємно посилюючи специфічні ефекти (потенціюють експресію гена).
Кількість рецепторів клітинної мембрани і внутрішньоклітинних рецепторів змінюється, змінюється і сила їх зв'язку з гормоном. Клітини міометрія і молочних залоз містять окситоциновий рецептори, число яких зростає під дією естрогенів (up-регулювання) і знижується під дією прогестерону (down-регулювання). У міокарді містяться норадреналінового рецептори (β 1), число яких і спорідненість до норадреналіну підвищується під дією тиреоїдних гормонів (Т3 / Т4).
Водорозчинні гормони (моноаміни, амінокислоти і пептиди) зв'язуються з рецепторами мембрани, яка насичена ліпідами, тому не дозволяє вільно дифундувати водорозчинних гормонів через мембрану. В гормональної реакції клітини водорозчинні гормони називають першими мессенджерами. У відповідь на їх взаємодію з рецептором всередині клітини активуються так звані другі месенджери - цАМФ, циклічний гуанозинмонофосфат, інозітолтріфосфат, іони кальцію, диацилглицерол і ін. Іони кальцію служать дуже важливим другим месенджером. Потік іонів кальцію через клітинну мембрану в цитозоль контролюється гормон-рецептор-ної зв'язком, нервовими стимулами або модифікується іншими другими мессенджерами.
Концентрація гормонів в більшості випадків становить 10 -10 моль / л. При цьому зв'язування однієї молекули з мембранним рецептором призводить до утворення 10 000 молекул цАМФ в клітці, і в цьому відношенні цАМФ виступає в якості молекулярного підсилювача гормонального сигналу (в 10000 разів!). Фосфодіестерази руйнує цАМФ, тому її інгібітори - теофілін та кофеїн - діють синергічно з гормонами, у яких цАМФ є другим месенджером. цАМФ стимулює катаболические процеси - ліполіз, глікогеноліз (глюкагон), глюконеогенез і кетогенез, секрецію інсуліну β-клітин і підшлункової залози.
Реферат на тему
Ендокринні залози і їх гормони
ендокринні залози
(Від грец. Endon - всередині, crio - виділяю) або залози внутрішньої секреції, є спеціалізовані органи або групи клітин, основна функція яких полягає у виробленні та виділення у внутрішню середу організму специфічних біологічно активних речовин. Залози внутрішньої секреції не мають вивідних проток. Їх клітини обплетені рясної мережею кровоносних і лімфатичних судин, і продукти життєдіяльності виділяються безпосередньо в кров, лімфу, тканинну рідину. Ця особливість принципово відрізняє ендокринні залози від екзокринних, які виділяють свої секрети через вивідні протоки. Продукти, що виробляються залозами внутрішньої секреції, отримали назву гормонів(Грец. Hormao - збуджую, активують). Термін «внутрішня секреція» був запропонований в 1885 році французьким фізіологом К. Бернаром, а термін «гормон» - англійськими фізіологами У. Бейлісом і Е. Старлінг в 1902 році.
Для гормонів характерні наступні особливості: Синтез і виділення їх здійснюється спеціалізованими клітинами.
Гормони утворюються в залізистих ендокринних клітинах, після чого вони надходять у внутрішнє середовище, в основному в кров і лімфу. Проміжні продукти синтезу або метаболізму гормонів нерідко мають біологічну активність, але, як правило, не серкетіруются .
Висока біологічна активність.
Гормони роблять фізіологічна дія в дуже малих концентраціях. Так, концентрація жіночого статевого гормону (естрадіолу) в крові коливається від 0,2 до 0,6 мкг (Ю -6 г) в 100 мл плазми. Зміст гормону росту в крові вимірюється ще меншими величинами - наніграммамі (КГ 9 г). Гіпофіз реагує на пікограм (Ю -12 г) гіпоталамічних гормонів, ангіотензин-П-продукт ендокринних клітин нирок - викликає відчуття спраги в фемтограмм (Ю -15 г). Крім гормонів, ніякі інші хімічні продукти життєдіяльності не ефективні в таких малих дозах. Специфічність.Кожен гормон характеризується певними, притаманними тільки йому хімічною структурою, місцем синтезу і функцією. У зв'язку з цим дефіцит будь-якого гормону не може бути заповнений іншими гормонами або біологічно активними речовинами. Дистантность дії.
Гормони, як правило, переносяться кров'ю далеко від місця освіти, впливаючи на віддалені органи і тканини. Цим вони відрізняються від медіаторів і цитокінів, що діють на одну клітку або групу клітин на місці їх утворення.
Хімічна структура гормонів і їх перетворення в організмі
За хімічною будовою гормони можна розділити на 4 основні групи:
білки і пептиди;
похідні амінокислот;
стероїди;
простагландини.
Прикладами білкових гормонів можуть служити інсулін, соматотропін (гормон росту), гормони тропів передньої долі гіпофіза. Деякі з них (фоллітропін, тиреотропин, лютропін) відносяться до складних білок, інші (інсулін, кальцитонін та ін.) - до простих білків. Пептидную структуру мають глюкагон, вазопресин, окситоцин, гіпоталамічні гормони. До похідних амінокислот відносяться гормони щитовидної залози - трийодтиронін, тироксин, а також адреналін і норадреналін. Стероїдні гормони мають в своїй основі ядро \u200b\u200bциклічного вуглеводню циклопентанпергідрофенантрену. До цієї групи належать гормони кори надниркових і статевих залоз. Основні етапи освіти і перетворення гормону можна представити таким чином:
біосинтез гормону;
секреція, тобто виділення з ендокринної клітини;
транспорт кров'ю до периферичних тканин;
розпізнавання гормонального сигналу клітинами-мішенями;
трансдукция (переклад) гормонального сигналу в біологічний відповідь;
гасіння гормонального сигналу.
Органи і тканини з инкреторной функцією клітин
№
|
Органи і тканини
|
клітини
|
гормони
|
1.
|
плацента
|
синцитіотрофобласти цитотрофобласт
|
Хоріонічний гонадотропін, пролактин Естріол, прогестерон
|
2.
|
тимус
|
тимоцити
|
Тимозин, тимопоетин
|
3.
|
нирка
|
ПІВДНЯ інтерстицій
канальці
|
ренін еритропоетин
кальцитриол
|
4.
|
серце
|
міоцити передсердь
|
Атріопептід, соматостатин, ангіотензин-П
|
Органи, тканини і клітини з ендокринною функцією
№
|
структури
|
Тканини, клітини
|
гормони
|
1.
|
гіпоталамус
|
Нервові клітини мелкоточечних структур
|
Гіпоталамічні нейрогормони:
|
|
|
|
заднього гіпоталамуса Нервові клітини крупноклітинних ядер переднього гіпоталамуса
|
а) ліберіни: кортиколиберин, тіроліберін, люліберін, фолліберін, соматолиберин, пролактоліберін, меланоліберін; б) статини: пролактостатін, меланостатин, соматостатин
Вазопресин, окситоцин
|
2.
|
гіпофіз аденогіпофіз нейрогіпофіз
|
Кортікотрофи гонадотрофов
тіреотрофов
соматотрофов
лактотрофи
Пітуіціти
|
Кортикотропін, меланотро- пін Фоллітропін, лютропін Тиреотропін Соматотропин Пролактин
Вазопресин, окситоцин
|
3.
|
наднирники Корковаречовина
мозкова речовина
|
Клубочковая зона Пучкова зона Сітчаста зона Хромафині клітини
|
Мінералокортикоїди Глюкокортикоїди Статеві стероїди: андрогени, естрогени Андреналін, норадреналін
|
4.
|
Щитовидна залоза
|
фолікулярні тиреоцитах К-клітини
|
Трийодтиронин, тетрайодтіронін Кальцитонин, катакальцін, Ко-кальцігенін
|
5.
|
околощітовідние залози
|
Головні клітини К-клітини
|
паратирин Кальцитонин
|
6.
|
епіфіз
|
Пінеоціти
|
мелатонін
|
7.
|
Підшлункова залоза
|
Острівці Лангерганса: ос-клітини Р-клітини А-клітини
|
глюкагон інсулін
соматостатін
|
8.
|
Статеві залози (насінники, яєчники)
|
Клітини Лейдіга клітини Сертолі клітини гранульози жовте тіло
|
Тестостерон Естроген, ингибин Естрадіол, естрон, прогестерон прогестерон
|
біосинтез гормонівзапрограмований в генетичному апараті спеціалізованих ендокринних клітин. Отже, він залежить від структури і експресії генів, що кодують синтез цих гормонів, а також від ферментів, що регулюють синтез гормону і Посттрансляційні процеси. Відсутність або дефект відповідних генів призводить до ендокринопатії. Прикладом може служити карликовість при генетичному дефекті гормону росту.
2.Секреція гормонів.Як вказувалося, найважливіша ознака будь-якого гормону - його секретируемой. Упакований в везикули або гранули гормон транспортується у напрямку до цитоплазматичної мембрани. Виходу білково-пептидних гормонів і катехоламінів з клітки передує взаємодія цитоплазматичної мембрани і мембрани секреторною гранули. Після цього відбувається їх лізис і вихід гормону з клітки. Даний процес активується багатьма факторами медіаторами, високою концентрацією калію, електричними стимулами і ін. Секреція гормону - акт, супроводжуваний витратою енергії, тому вона завжди пов'язана зі зрушеннями в системі АТФ - цАМФ. Для секреції необхідна участь іонів кальцію, які активують білки мікротубулярної-мікрофіламентною системи, сприяючи взаємодії з ними гранул гормону, і впливають на утворення цАМФ. Тому зниження вмісту іонів кальцію в позаклітинній середовищі і надходження його в ендокринну клітку неминуче призводить до зменшення секреторної активності цієї клітини. 3.
Транспорт гормонів.Секретується гормон потрапляє у внутрішнє середовище організму, переважно в кров, і транспортується далі. Більшість гормонів утворює в крові комплексні сполуки з білками плазми. Частина цих білків - специфічні транспортні протеїни (наприклад, транскортином, що зв'язує гормони кори надниркових залоз), частина - неспецифічні (наприклад, у-глобуліни). Комплексирование з білками - оборотний процес. Крім того, частина гормонів пов'язана в крові з форменими елементами, зокрема еритроцитами. Освіта пов'язаної форми гормонів має велике фізіологічне значення. По-перше, це оберігає організм від надмірного накопичення в крові (і, отже, впливу на тканини) вільних гормонів. По-друге, пов'язана форма гормону є його фізіологічним резервом. По-третє, зв'язування з білками сприяє захисту гормону від руйнування ферментами, тобто продовжує його життя. Нарешті, комплексування з білками перешкоджає фільтрації мелкомолекулярних гормонів через ниркові клубочки і тим самим утримує ці важливі регуляторні процеси.
4.
Розпізнавання гормонального сигналу.Після надходження в периферичні органи гормони, як правило, звільняються від білкового компонента і, фіксуючи на певних рецепторах клітин, що сприймають цей гормон (клітинах-мішенях), здійснюють своє специфічне дію. В процесі периферичної дії гормонів відбуваються їх різні перетворення. При цьому можливе утворення нових гормональних продуктів, нерідко більш активних або виробляють інше біологічне дію, ніж вихідний гормон. Так, гормон щитовидної залози тироксин може в тканинах перетворюватися в трийодтиронін - більш активний гормон цієї ж залози. Андрогени (чоловічі статеві гормони) перетворюються в гіпоталамусі в естрогени - жіночі статеві гормони. Трансдукція гормонального сигналу в біологічний відповідь органічно пов'язана з механізмом дії цього гормону. Дія будь-якого гормону на клітини-мішені завжди починається з взаємодії його з певними компонентами клітини. Це явище називається рецепцією гормону, а клітинні компоненти, які взаємодіють з гормоном, - рецепторами. Рецепторами гормонів є кислі крупномолекулярні олігопептиди.
Структура рецепторной молекули характеризується асиметричністю. Виділяють три її ділянки:
зв'язує гормон;
еффекторний, передає гормональний сигнал на внутрішньоклітинні механізми, тобто здійснює трансдукцію сигналу в біологічний відповідь;
сполучають перший і другий ділянки.
Рецептори розташовуються або внутрішньоклітинно, або на поверхні цитоплазматичної мембрани. Внутрішньоклітинні рецептори пов'язують стероїдні і тиреоїдні гормони, мембранні рецептори - інші гормони. Вважається також, що функція розпізнавання специфічного гормонального сигналу у всіх клітин для всіх гормонів здійснюється мембранним рецептором, а після зв'язування гормону з відповідним йому рецептором подальша роль гормон-рецепторного комплексу для пептидних та стероїдних гормонів різна. У пептидних, білкових гормонів і катехоламінів гормон-рецепторний комплекс призводить до активації мембранних ферментів і утворення різних вторинних посередників (месенджерів) гормонального регуляторного ефекту, що реалізують свою дію в цитоплазмі, органелах і ядрі клітини.
Відомі чотири системи вторинних посередників:
аденилатциклаза - циклічний аденозинмонофосфат (цАМФ);
гуанілатциклазу - циклічний гуанозинмонофосфат (цГМФ);
фосфоліпаза С - інозітолтріфосфат (ІТФ);
іонізований кальцій.
4.1.
Система «аденилатциклаза - цАМФ».
Мембрани фермент аденилатциклаза може перебувати в двох формах - активованої і неактивованої. Активація аденілатциклази відбувається під впливом гормон-рецепторного комплексу, утворення якого призводить до зв'язування гуанілова нуклеотиду (ГТФ) з особливим регуляторним стимулюючим білком (GS-білок), після чого GS-білок викликає приєднання магнію до аденілатциклази і її активацію. Так діють активізують аденілатциклазу гормони глюкагон, тиреотропин, паратирин, вазопресин, гонадотропін та ін. Деякі гормони, навпаки, пригнічують аденілатциклазу (соматостатин, ангіотензин-П та ін.). Під впливом аденілатциклази з АТФ синтезується цАМФ, викликає активацію протеїнкінази в цитоплазмі клітини, що забезпечують фосфорилювання численних внутрішньоклітинних білків. Це змінює проникність мембран, тобто викликає типові для гормону метаболічні і, відповідно, функціональні зрушення. Внутрішньоклітинні ефекти цАМФ проявляються також у впливі на процеси проліферації, диференціювання, на доступність мембранних рецепторних білків молекулам гормонів.
Система «гуанілатциклазу - цГМФ».
Активація мембранної гуанілатциклази відбувається не під безпосереднім впливом гормон-рецепторного комплексу, а опосередковано через іонізований кальцій і оксидантного системи мембран. Так реалізують свої ефекти натрийуретический гормон передсердь - атріопептід, тканинної гормон судинної стінки. У більшості тканин біохімічні та фізіологічні ефекти цАМФ і цГМФ протилежні. Прикладами можуть служити стимуляція скорочень серця під впливом цАМФ і гальмування їх цГМФ, стимуляція скорочень гладких м'язів кишечника цГМФ і придушення цАМФ.
4.3 Система «фосфоліпаза С - ііозітолтріфосфат».
Гормон-рецепторний комплекс за участю регуляторного білка G веде до активації мембранного ферменту фосфоліпази С, що викликає гідроліз фосфоліпідів мембрани з утворенням двох вторинних посередників - інозітолфосфата і діацілгліцерола. Ііозітолтріфосфат викликає вихід іонізованого кальцію з внутрішньоклітинних депо. Останній, зв'язуючись з білком кальмодулином, забезпечує активацію протеїнкінази і фосфорилювання внутрішньоклітинних структурних білків і ферментів. Диацилглицерол також активує і завершує процес фосфорилювання інших білків, одночасно реалізуючи і другий шлях гормонального ефекту: через утворення арахідонової кислоти, що є джерелом потужних по метаболічним і фізіологічним ефектам речовин - простагландинів і лейкотрієнів. Через розглянуті системи вторинних посередників реалізуються ефекти адреналіну, вазопресину, ангіотензину-П, соматостатину, окситоцину і деяких інших гормонів.
4.4. Система «кальцій - кальмодулін».
Іонізований кальцій надходить в клітку після утворення гормон-рецепторного комплексу або з позаклітинного середовища за рахунок активування повільних кальцієвих каналів (наприклад, в міокарді), або з внутрішньоклітинних депо під впливом вищеописаних внутрішньоклітинних процесів. У цитоплазмі нем'язові клітин кальцій зв'язується зі спеціальним білком - кальмодулином, а в м'язових клітинах роль кальмодулина виконує тропонин С. Пов'язаний з кальцієм кальмодулін активує численні протеїнкінази, що забезпечують фосфорилювання білків. Короткочасне збільшення в клітці кількості кальцію і його зв'язування з кальмодулином є стимулом для численних фізіологічних процесів - скорочення м'язів, секреції гормонів і виділення медіаторів, синтезу ДНК, зміни рухливості клітин, активності ферментів, транспорту речовин через мембрани. У стероїдних гормонів мембранний рецептор забезпечує специфічне впізнавання гормону і його перенесення в клітку. У цитоплазмі є особливий цитоплазматический білок-рецептор, з яким зв'язується гормон. Цей зв'язок з рецепторним білком необхідна для вступу стероїдного гормону в ядро, де відбувається його взаємодія з третім ядерним рецептором, зв'язування комплексу «гормон - ядерний рецептор» з хроматиновой акцептором, специфічним кислим білком і ДНК, що тягне за собою активацію транскрипції мРНК, синтез транспортних і рибосомних РНК, транспорт мРНК в цитоплазму, трансляцію мРНК з синтезом білків і ферментів в рибосомах. Всі ці явища вимагають тривалої присутності гормон-рецепторного комплексу в ядрі. Однак ефекти стероїдних гормонів проявляються не тільки через кілька годин, частина з них виникає швидко, протягом декількох хвилин. Це такі ефекти, як підвищення проникності мембран, посилення транспорту глюкози і амінокислот, звільнення лізосомальних ферментів, зрушення енергетики мітохондрій і, крім того, збільшення цАМФ і іонізованого кальцію. Таким чином, цілком обгрунтований погляд, прихильники якого вважають, що мембранний рецептор стероїдних гормонів не тільки виконує функцію «впізнавання» молекули гормону, але і, подібно до рецепторам пептидних гормонів, активує систему вторинних посередників в клітці. Пептидні гормони також мають здатність вибірково впливати на транскрипцію генів в ядрі клітини. Цей ефект може бути реалізований не тільки з поверхні клітини за рахунок вторинних посередників, а й шляхом надходження гормонів всередину клітини за рахунок інтернетизаціїгормон-рецепторного комплексу (завдяки ендоцитозу).
Феномен інтерналізації гормон-рецепторних комплексів і зменшення тим самим кількості рецепторів до гормону на мембрані клітини дозволяє зрозуміти механізм зниження чутливості ефектора при надмірній кількості гормональних молекул, або феномен десенситизацииефектора. Це явище, по суті, є негативним зворотним регуляторної зв'язком на рівні ефектора. Протилежне явище - сенситизация,або підвищення чутливості до гормонів, може бути обумовлено збільшенням числа вільних рецепторних місць на мембрані як за рахунок падіння інтерналізації, так і в результаті «спливання» активних зв'язуючих ділянок рецепторів. Таким чином, гормони передають клітці інформаційні сигнали, а сама клітина здатна регулювати ступінь сприйняття гормонального контролю. 5. Трансдукція гормонального сигналу в біологічний відповідь.Розрізняють 5 видів дії гормонів на тканини-мішені: метаболічну, морфогенетическое, кінетичне, коригуючий, реактогенність. Метаболічну дію.
Зрушення метаболізму, викликані гормонами, лежать в основі зміни функції клітин, тканини і органу. Морфогенетическое дію -
вплив гормонів на процеси формоутворення, диференціювання і зростання структурних елементів. Прикладами можуть служити вплив соматотропіну на зростання тіла і внутрішніх органів, вплив статевих гормонів на розвиток вторинних статевих ознак. кінетичне дію
- здатність гормонів запускати діяльність ефектора, включати реалізацію певної функції. Наприклад, окситоцин викликає скорочення мускулатури матки, адреналін запускає механізм розпаду глікогену в печінці і вихід глюкози в кров, визопрессін включає механізм зворотного всмоктування води в збірних трубочках нефронів. коригуючий дію
- зміна діяльності органів або процесів, які відбуваються і під час відсутності гормону. Прикладами коригуючого дії гормонів є вплив адреналіну на частоту серцевих скорочень, активація окислювальних процесів тироксином, зменшення зворотного всмоктування іонів калію в нирках під впливом альдостерону. Різновидом коригуючого дії є нормалізує ефект гормонів, коли їх вплив направлено на відновлення зміненого або навіть порушеного процесу. Наприклад, при вихідному превалювання анаболітіческое процесів білкового обміну глюкокортикоїди викликають катаболический ефект, але якщо початково переважає розпад білків, глюкокортикоїди стимулюють їх синтез. Реактогенпое дію
- здатність гормону міняти реактивність тканини до дії того ж гормону, інших гормонів або медіаторів нервових імпульсів. Так, наприклад, тиреоїднігормони посилюють дію катехоламінів, кальцій-регулюючі гормони знижують чутливість дистальних відділів нефрона до дії вазопресину. Різновидом реактогенність дії гормонів є пермісивними дію, яка передбачає здатність одного гормону давати можливість реалізуватися ефекту іншого гормону. Так, наприклад, глюкокортикоїди володіють пермісивними дією по відношенню до катехоламінів (для реалізації ефектів адреналіну необхідна присутність малих кількостей кортизолу); інсулін володіє пермісивними дією по відношенню до соматотропіну і т.д. 6. Гасіння гормонального сигналу.Даний механізм діє з метою нормалізації гормонального сигналу (при його затримці возникет небезпека надмірної напруги багатьох функцій). Існують численні механізми гасіння гормонального сигналу. Перш за все, це зменшення биосинтетической і секреторної активності самих гормон-продукують клітин. Поряд з цим відбувається руйнування (катаболізм) частини виділених гормонів. В результаті значна частка гормонів зникає з крові після того, як вони вивільнилися з гранул. Ферменти печінки інактивують гормони шляхом дезамінування і метилування. У процесі інактивації як гормони, так і продукти їх катаболізму кан'югіруются в печінці з глюкуроновою і сірчаною кислотами, що збільшує їх водорастворимость і полегшує подальше виділення через нирки. Частина вільних і кон'югованих гормонів надходить в жовч і виводиться з організму через кишечник. Гасіння гормонального сигналу сприяє описана вище десенситизация тканин при тривало підвищеному рівні гормону в крові. У ряді випадків утворюються антитіла до гормонів і інші інгібітори, які гальмують біологічний ефект.
Зниження активності гормонального сигналу досягається і в результаті секреції контррегуляторних гормонів, тобто гормонів протилежної дії. Наприклад, надлишок інсуліну викликає гіпоглікемію, яка, в свою чергу, індукує секрецію контрінсулярних гормонів. Ще одним важливим фізіологічним механізмом гасіння гормонального сигналу є гальмування секреції за принципом зворотного зв'язку.
Сучасне вчення про регуляторних пептидах
регуляторні пептиди- біологічно активні речовини, що синтезуються різними за походженням клітинами організму і беруть участь в регуляції різних функцій. Серед них виділяють нейропептиди, які секретуються нервовими клітинами і беруть участь в здійсненні функцій нервової системи. Крім цього, вони виявлені і за межами ЦНС в ряді ендокринних залоз, а також в інших органах і тканинах.
В онтогенезі регуляторні пептиди з'явилися значно раніше «класичних» гормонів, тобто до відокремлення спеціалізованих ендокринних залоз. Це дозволяє вважати, що в геномі запрограмовано роздільне освіту названих груп речовин, а отже вони є самостійними.
Джерелами регуляторних пептидів служать поодинокі гормон-продукують клітини, що утворюють іноді невеликі скупчення. Ці клітини розглядають як початкову форму ендокринних утворень. До них відносяться нейросекреторні клітини гіпоталамуса, нейроендокринні (хромафінні) клітини наднирників і параганглиев, клітини слизової оболонки гастро-інтестинального системи, пінеалоціти епіфіза. Встановлено, що ці клітини здатні декарбоксилировать ароматичні кислоти-попередники нейроамінов, що дозволило об'єднати їх в єдину систему (Pearse, 1976), що отримала назву «APUD-система» (за першими літерами англійсікх слів Amine Precursor Uptake and Decarboxylating system - система захоплення і декарбоксилювання попередників амінів). Велике число пептидів (вазоактивний інтестинального пептид - ВІП, холецистокінін, гастрин, глюкагон) спочатку було виявлено в секреторних елементах гастро-інтестинального тракту. Інші (субстанція Р, нейротензин, енкефаліни, соматостатин) були спочатку знайдені в нервовій тканині. Слід зазначити, що в гастро-інтестинального тракту деякі пептиди (гастрин, холецистокінін, ВІП і деякі інші) присутні і в нервах, а також і в ендокринних клітинах.
Існування цієї нейродіффузной ендокринної системи пояснюють міграцією клітин з єдиного джерела - нервового гребінця; вони включаються в ЦНС і в тканини різних органів, де перетворюються в ЦНС-подібні Кетков, секретуючі нейроаміни (нейромедіатори) і пептидні гормони. Це пояснює присутність нейропептидів в кишечнику і підшлунковій залозі, клітин Кульчицького в бронхах, а також робить зрозумілим виникнення гормонально-активних пухлин легенів, кишечника, підшлункової залози. Апудоціти зустрічаються також в нирках, серце, лімфатичних вузлах, кістковому мозку, епіфізі, плаценті.
Основні групи регуляторних пептидів (по Krieger)
Найбільш поширеною є класифікація регуляторних пептидів, що включає наступні групи:
гипоталамические рилізинг-гормони;
нейрогіпофізарние гормони;
пептиди гіпофіза (АКТГ, МСГ, СТГ, ТТГ, пролактин, ЛГ, ФСГ, (3-ендорфін, липотропини);
гастро-інтестинального пептиди;
інші пептиди (ангіотензин, кальцитонін, нейропептид V).
Для ряду пептидів встановлені локалізація містять і клітин і розподіл волокон. Описано кілька пептідергіческіх систем мозку, які поділяють на два основних види.
Довгі проекційні системи,
волокна яких досягають віддалених областей мозку. Наприклад, тіла нейронів сімейства проопиомеланокортина розташовані в аркуатних ядрі гіпоталамуса, а їх волокна досягають мигдалини і околоводопроводного сірої речовини середнього мозку.
Короткі проекційні системи:
тіла нейронів розташовані нерідко в багатьох областях мозку і мають локальний розподіл відростків (субстанція Р, енкефаліни, холецистокінін, соматостатин).
Багато пептиди присутні в периферичних нервах. Наприклад, субстанція Р, ВІП, енкефаліни, холецистокінін, соматостатин виявлені в блукаючого, чревного і сідничного нервах. Мозкова речовина надниркових залоз містить велику кількість препроенкефаліна А (метенкефалін). Показано існування нейропептидів і нейротрансмітерів в одному і тому ж нейроне: серотонін виявлений в нейронах довгастого мозку разом з речовиною Р, допамін разом з холецистокинином - в нейронах середнього мозку, ацетилхолін і ВІП - в вегетативних гангліях. Про функціональне значення цього співіснування дозволяють судити такі фактори. Під впливом ВІП в фізіологічних концентраціях відбувається виражене збільшення чутливості до ацетилхоліну мускаринових рецепторів в підщелепної залозі котів, а антисироватка до ВІП частково блокує вазодилатацію, викликану стимуляцією парасимпатичних нервів.
Синтез регуляторних пептидів.
Характерною особливістю синтезу пептидів є їх освіту шляхом фрагментації великої молекули-попередника, тобто в результаті так званого посттрансляційних протеолітичного розщеплення - процесингу. Синтез попередника відбувається в рибосомах, що підтверджується наявністю матричної РНК, що кодує пептид, а Посттрансляційні ензимні модифікації з виділенням активних пептидів - в апараті Гольджі. Ці пептиди досягають нервових закінчень завдяки аксонального транспорту.
Активні пептиди, що походять з одного попередника, утворюють його сімейство. Описано наступні сімейства пептидів.
Сімейство проопиомеланокортина (ПОМК).Тіла нейронів, в яких присутній цей великий білок (286 амінокислотних залишків), локалізуються в аркуатних ядрі гіпоталамуса. Залежно від набору ферментів з ПОМК утворюються: у передній долі гіпофіза - переважно АКТГ, (3-ліпотропін, Р-ендорфін, в проміжній - сх-меланостимулирующего гормон і Р-ендорфін. Таким чином, набір ферментів визначає спеціалізацію продукції клітинами суворо визначених пеп - тідов. Це ферменти катепсини В, трипсин, карбоксипептидаза, амінопептідазу, місця їх атаки - парні залишки амінокислот.
Сімейство церулеіна:гастрин, холецистокінін.
Сімейство ВІП: секретин, глюкагон.
Сімейство аргінін-вазопресину:вазопресин, окситоцин.
Крім того, встановлено, що мет-енкефалінів і лей-енкефалінів мають попередників у вигляді препроенкефаліна А і препроенкефаліна В відповідно. Протеолиз в даному випадку - НЕ інактивація, а трансформація активності. Механізм дії нейропептидів:
Характерною особливістю регуляторних пептидів є поліфункціональність (за механізмом і характером ефектів) і освіту регуляторних ланцюгів (каскадів). В цілому механізми дії пептидів можна розділити на дві групи: синаптичні і внесінаптіческого. 1.
Синаптичні механізми дії пептидівможуть виражатися в нейромедіаторної або нейромодуляторной функції. Нейромедіатор (пейротрансміттер)- речовина, що вивільняється з пресинаптичної терміналі і діє на наступну - постсинаптическую мембрану, тобто виконує передавальну функцію. Встановлено, що деякі пептиди виконують цю функцію через пептідергіческіе рецептори, наявні на нейронах (їх тілах або терміналах). Так, гіпоталамічний релизинг-гормон лютеїнізуючого гормону (люліберін) в синаптичних гангліях жаби виділяється при стимуляції нерва за допомогою кальцій-залежного процесу і викликає пізній повільний збудливий постсинаптичний потенціал.
На відміну від «класичних» нейротрансмітерів (норадреналіну, допаміну, серотоніну, ацетилхоліну), пептиди, які виконують передавальну функцію, характеризуються високою аффинностью рецепторів (що може забезпечити більш дистантное дію) і тривалим (десятки секунд) дією в зв'язку з відсутністю ферментних систем інактивації і зворотного депонування.
нейромодуляторов,на відміну від нейротрансмітера, не викликає самостійного фізіологічного ефекту в постсинаптичні мембрані, але модифікує реакцію клітини на нейромедіатор. Таким чином, нейромодуляціі - НЕ передавальна, а регуляторна функція, яка може здійснюватися як на пост-, так і на пресинаптичних рівні. Види нейромодуляціі:
контроль виділення нейротрансмітера з терминалей;
регуляція кругообігу нейротрансмітера;
модифікація ефекту «класичного» нейротрансмітера.
2.
Внесінаптіческого дію пептидівреалізується декількома шляхами. А. паракрінним дію(Паракрінним) - здійснюється в зонах міжклітинної контакту. Наприклад, соматостатин, що виділяється А-клітинами острівцевих тканини підшлункової залози, виконує паракринного функцію в контролі секреції інсуліну і глюкагону (3- і ос-клітинами відповідно, а кальцитонін - в контролі секреції йодовмісних гормонів щитовидною залозою. Б. нейроендокринних дію- здійснюється через виділення пептиду в кров'яне русло і його вплив на клітину-ефектор. Прикладами можуть служити соматостатин і інші гіпоталамічні фактори, які виділяються в медіальній еміненції з деяких терминалей в портальний кровотік і контролюючі секрецію гіпофізарних гормонів. В. Ендокринна дію.В даному випадку пептиди виділяються в загальний кровотік і діють як дистантних регулятори. Цей механізм включає компоненти, обов'язкові для «класичних» ендокринних функцій, - транспортні білки і рецептори клітин-мішеней. Так встановлено, що в якості переносників-стабілізаторів використовуються: нейрофізін - для вазопресину і окситоцину, деякі альбуміни і глобуліни плазми - для холецистокініну і гастрину. Що стосується рецепції, то існування відокремлених рецепторів встановлено для опіоїдних пептидів, вазопресину, ВІЛ. В якості вторинних месенджерів можуть використовуватися циклічні нуклеотиди, продукти гідролізу фосфоінозитидів, кальцій і кальмодулін з наступною активацією протеїнкінази і контролем фосфорилювання білків-регуляторів трансляції та транскрипції. Крім того, описаний механізм інтерналізації, коли регуляторний пептид разом з рецептором проникає в клітину за допомогою механізму, близького до піноцитозу, і відбувається передача сигналу в геном нейрона. Для регуляторних пептидів характерне утворення складних ланцюгів або каскадів в результаті того, що утворюються з основного пептиду метаболіти теж функціонально активні. Цим пояснюють тривалість ефектів короткоживучих пептидів.
Функції регуляторних пептидів: 1. Біль.Цілий ряд пептидів впливає на формування болю як складного психофізіологічного стану організму, що включає саме болюче відчуття, а також емоційні, вольові, рухові і вегетативні компоненти. При цьому пептиди включені як в ноцицептивную, так і в антиноцицептивную систему. Так, речовина Р, соматостатин, ВІП, холецистокінін і ангіотензин виявлені в первинних сенсорних нейронах, причому речовина Р є нейротрансмиттером, які виділяються певними класами аферентних нейронів. У той же час, енкефаліни, вазопресин, ангіотензин і родинні опіоїдні пептиди виявлені в низхідному супраспинального шляху, що йде до задніх рогів спинного мозку і яке справляє гальмівну дію на ноцицептивних шляху (аналгетичний ефект).
2. Пам'ять, навчання, поведінку.Отримано дані про те, що фрагменти АКТГ (АКТГ 4-7 і АКТГ 4-10), позбавлені гормональних ефектів, і сс-меланостимулирующего гормон покращують короткочасну пам'ять, а вазопресин залучений у формування довготривалої пам'яті. Введення в мозкові шлуночки антитіл до вазопресину протягом години після сеансу навчання викликає забування. Крім того, АКТГ 4-10 покращує увагу.
Встановлено вплив ряду пептидів на харчову поведінку. Прикладами можуть служити посилення харчової мотивації під дією опіоїдних пептидів і ослаблення - під дією холецистокініну, кальцитоніну і кортиколиберина.
Опіоїдні пептиди роблять значний вплив на емоційні реакції, будучи ендогенними ейфоригенним.
ВІП надає снодійну, гіпотензивну і бронхолітичну дію. Тиреоліберином дає псіхотонізірующій ефект. Люліберін, крім виконання командної функції (стимуляція гонадотропи передньої долі гіпофіза), регулює статеве і батьківське поводження.
3. Вегетативні функції.Цілий ряд пептидів бере участь в контролі рівня артеріального тиску. Це ренін-ангіотензинової системи, всі компоненти якої присутні в мозку, опіоїдні пептиди, ВІП, кальцитонін, атріопептід, що володіють сильним натрійуретічеським ефектом.
Описано зміни терморегуляції під дією деяких пептидів. Так, внутріцентральних введення тиреоліберином і Рендорфіну викликає гіпертермію, в той час як введення АКТГ і ос-МСГ - гіпотермію.
4. Стрес.Заслуговує великої уваги той факт, що ряд нейропептидів (опіоїдні пептиди, пролактин, пептиди епіфіза) відносять до антистресовий системі, оскільки вони обмежують розвиток стресових реакцій. Так, в експериментах з різними моделями показано, що опіоїдні пептиди обмежують активацію симпатичного відділу нервової системи і всіх ланок гіпоталамо-гіпофізарно-адреналової системи, попереджаючи виснаження цих систем, а також небажані наслідки надлишку глюкокортикоїдів (пригнічення запальної реакції і тимико-лімфатичної системи, поява виразок шлунково-кишкового тракту та ін.) - Антігіпоталаміческіе чинники епіфіза гальмують утворення либеринов і секрецію гормонів передньої долі гіпофіза. Зниження активації гіпоталамуса обмежує гиперсекрецию вазопресину, який надає шкідливу дію на міокард.
5. Вплив на імунну систему.Встановлено двосторонні зв'язки між системою регуляторних пептидів і імунною системою. З одного боку, в даний час достатньо вивчена здатність багатьох пептидів модулювати імунні відповіді. Відомі супрессия синтезу імуноглобулінів під дією (З-ендорфіну, енкефалінів, АКТГ і кортизолу; пригнічення секреції інтерлейкіну -1
(ІЛ -1)
і розвиток лихоманки під впливом а-меланоцітстімулірующего гормону. Встановлено, що вазоактивний інтестинального пептид (ВІЛ) гальмує всі функції лімфоцитів і їх вихід з лімфовузлів, що розцінюється як нова форма иммуномодуляции. У той же час, цілий ряд пептидів надає стимулюючу дію на імунну систему, викликаючи збільшення синтезу імуноглобулінів і у-інтерферону (| 3-ендорфін, тиреотропний гормон), посилення активності природних клітин-кілерів (Р-ендорфін, енкефаліни), збільшення проліферації лімфоцитів і виділення лімфокінів (субстанція Р, пролактин, гормон росту), підвищення продукції супероксидних аніонів (гормон росту). Описано рецептори лімфоцитів до ряду гормонів. З іншого боку, іммуномедіатори впливають на обмін і виділення гіпоталамічних нейротрансмітерів і рилізинг-гормонів. Так, регуляторний лейкопептід ІЛ -1
здатний проникати в мозок через ділянки підвищеної проникності гематоенцефалічний бар'єр і стимулювати секрецію кортикотропин-рилізинг-гормону (в присутності простагландину) з подальшою стимуляцією виділення АКТГ і кортизолу, які гальмують утворення ІЛ -1
і імунну відповідь.
Одночасно, через виділення соматостатина, ІЛ -1
пригнічує секрецію ТТГ і гормону росту. Таким чином, іммунопептід виконує роль тригера, який, замикаючи механізм зворотного зв'язку, попереджає надмірність імунної відповіді.
Відповідно до сучасних уявлень, повний регуляторний коло між нейроендокринними і імунними механізмами включає також пептиди, загальні для обох систем. Зокрема, показана здатність гіпоталамічних нейронів секретировать ІЛ-1. Виділено відповідальний за його продукцію ген, експресія якого індукується бактеріальними антигенами і кортикотропіном. Описано нейрональні шляху в медіобазальний гіпоталамус людини і щура, що містять ІЛ-1 і ІЛ-6, а також гіпофізарним клітини, що виділяють ці пептиди.
Таким чином, іммуномедіатори можуть регулювати функції передньої долі гіпофіза через:
ендокринний механізм (циркулюючі в крові лімфокіни активованих лімфоцитів);
нейроендокринні ефекти, реалізовані інтерлейкіну гіпоталамуса через тубероінфундибулярному портальну систему;
паракрінний контроль в самому гіпофізі.
З іншого боку, результати иммунохимических і молекулярних досліджень показали, що імунокомпетентні клітини секретують багато пептиди і гормони, пов'язані з ендокринною та нейрональної активністю: лімфоцити і макрофаги синтезують АКТГ; лімфоцити - гормон росту, пролактин, ТТГ, енкефаліни; мононуклеарние лімфоцити і огрядні клітини - ВІП, соматостатин; клітини тимуса - аргінін, вазопресин, окситоцин, нейрофізін. При цьому секретуються лімфоцитами гіпофізарні гормони регулюються тими ж факторами, що і гіпофіз. Наприклад, секреція АКТГ лімфоцитами пригнічується глюкокортикоїдами і стимулюється кортикотропин-рилізинг-гормоном. Запропоновано концепцію, згідно з якою виділення лімфоцитами перерахованих гормонів забезпечує аутокрінную і паракринного регуляцію локальної імунної реакції. Таким чином, функції трьох головних регуляторних систем - нервової, ендокринної та імунної - інтегровані в складні регуляторні кола, що функціонують за принципом зворотного зв'язку. При цьому периферичні лімфоцити, якщо слідувати концепції Д. Блелок (Blalock, 1989), забезпечують чутливий механізм, за допомогою якого розпізнаються некогнітівних стимули (чужорідні речовини) і мобілізуються нейроендокринні адаптивні відповіді.
Участь регуляторних пептидів у розвитку патології.
Оскільки пептидні гормони складають поліфункціональну систему, що бере участь в регуляції багатьох функцій в організмі, цілком ймовірно їх залучення в патогенез різних захворювань. Так, встановлено порушення концентрацій пептидів мозку при дегенеративних неврологічних захворюваннях невідомої етіології: хворобах Альцгеймера (зниження концентрації соматостатину в корі головного мозку) і Гантінгтона (зниження концентрації холецистокинина, речовини Р і енкефалінів, підвищення вмісту соматостатина в базальних гангліях, а також зменшення кількості рецепторів, зв'язують холецистокінін в цих структурах і в корі великих півкуль). Чи є ці зміни первинними або з'являються як наслідок розвитку захворювань, належить з'ясувати.
Відкриття опіоїдних пептидів і розподілу їх рецепторів в різних мозкових структурах, зокрема в лімбічної системі, привернуло увагу до оцінки їх значення в патогенезі психічних розладів. Запропоновано гіпотезу існування опіоїдної недостатності у хворих на шизофренію, зокрема неможливості освіти у-ендорфіну, що володіє нейролептическим дією. Встановлено збільшення концентрації атріопептід при застійних явищах в системі кровообігу, що, можливо, є механізмом компенсації порушень обміну натрію (його затримки).
Вивчення олігопептідних гормонів як регуляторної системи призвело до виділення особливої \u200b\u200bгрупи захворювань, обумовлених її патологією, - апудопатій.
Апудопатіі- захворювання, пов'язані з порушенням структури і функції апудоцитов і виражаються в певних клінічних синдромах. Розрізняють первинні апудопатіі, обумовлені патологією самих апудоцитов, і вторинні, що виникають як реакція апудоцитов на порушення гомеостазу організму, викликане захворюванням, патогенез якого первинно не пов'язаний з патологією APUD-системи (при інфекційних захворюваннях, пухлинному рості, хворобах нервової системи і т.д. ). Первинні апудопатіі можуть проявлятися в гіперфункції, гіпофункції, дисфункції, в освіті апудом - пухлин з клітин APUD-системи. Прикладами є наступні апудоми.
гастринома- апудома з клітин, що продукують гастрин, який, як відомо, стимулює виділення великої кількості шлункового соку з високою кислотністю і перетравлює силою. Тому клінічно гастринома проявляється розвитком ульцерогенного синдрому Золлінгера Еллісона. кортікотропінома- апудома, яка розвивається з апудобластов шлунково-кишкового тракту і виявляється ектопічної гиперпродукцией АКТГ і розвитком синдрому Іценко-Кушинга. Віпома- пухлина з клітин, які секретують вазоактивний інтестинального пептид. Локалізується в дванадцятипалій кишці або підшлунковій залозі. Виявляється розвитком водної діареї та зневодненням, а також розладом обміну електролітів. Соматостатинома- пухлина з клітин кишечника або острівцевих тканини підшлункової залози, які продукують соматостатин. Соматостатинома зазвичай розвивається як пухлина Д-клітин підшлункової залози, які секретують соматостатин. Характеризується клінічним синдромом, що включає цукровий діабет, жовчнокам'яну хворобу, гіпохлоргідрія, стеаторею і анемію. Діагностується з підвищення концентрації соматостатину в плазмі крові. Застосування регуляторних пептидів в медицині
На основі регуляторних пептидів створені деякі лікарські препарати. Так, олігопептиди (короткі пептиди) N-термінального фрагмента АКТГ і МСГ використовуються для корекції уваги і запам'ятовування, вазопресин - для поліпшення пам'яті при травматичної та інших амнезії. Широке застосування в лікувальній практиці має вітчизняний препарат даларгин (аналог лейенкефалін). Розпочато комерційний випуск сурфагона (аналог люліберіна), призначеного для корекції порушень репродуктивної системи. |