Як з'явилися птахи землі. Ці дивовижні древні птахи. Предок, якого може і не бути
Палеонтологічні матеріали з птахів, як зазначалося, дуже мізерні і уривчасті. Тому говорити про предків птахів серед рептилій і про походження та взаємини сучасних груп птахів доводиться переважно імовірно, ґрунтуючись на морфологічних особливостях сучасних груп і, дуже мало, на уривчастих палеонтологічних даних.
За сучасними уявленнями, предками птахів були архозаври Аrchosauria - велика і дуже різноманітна група рептилій, що панувала в мезозої і включала різних динозаврів, крокодилів, ящерів, що літають. Безпосередніх предків птахів, мабуть, потрібно шукати серед найдавнішої та примітивнішої групи архозаврів - текодонтів або псевдозухій Thecodontia (Pseudosuchia), що дали початок і решті більш високоорганізованих груп архозаврів. Псевдозухії на вигляд були схожі на ящірок. Вони вели переважно наземний спосіб життя, харчувалися, мабуть, різноманітною дрібною тваринною їжею. Задні ноги були трохи довші за передні, і при швидкому бігу тварини спиралися об грунт тільки задніми кінцівками. За багатьма морфологічними особливостями псевдозухії дуже подібні до птахів (деталі будови черепа, таза, задніх кінцівок). Проміжних форм, які б з'ясувати стадії розвитку оперення і вироблення здібності до польоту, доки знайдено. Вважають, що перехід до деревного способу життя і пристосування до стрибків з гілки на гілку супроводжувався розростанням і ускладненням структури рогових лусок на боках тулуба і хвоста, по задньому краю передніх і задніх кінцівок, що забезпечувало можливість планування та подовження стрибків (гіпотетична стадія Хейльманну, 1926, рис. 13).
Подальша спеціалізація та відбір призвели до розвитку оперення крила, що забезпечило можливість спочатку плануючого, а потім активного польоту. Оперення, що покриває все тіло птахів, ймовірно, розвивалося не так як пристосування до польоту (забезпечення обтічності тіла), як пристосування до термоізоляції; висловлюються припущення, що могло сформуватися ще до вироблення здатності до польоту. У деяких псевдозухій були подовжені рогові луски з чітким поздовжнім гребенем, від якого відходили дрібні поперечні реберця. Такі лусочки, певне, можна як структуру, з якої шляхом розчленування могло виникнути пташине перо.
Відокремлення птахів від плазунів, мабуть, відбулося вже наприкінці тріасу - початку Юри (170 - 190 млн. Років тому). Однак викопних залишків птахів цього часу поки що не знайдено. Досить хороші залишки двох найдавніших відомих нам птахів було знайдено у 60-х роках ХІХ ст. у Золенгофена (Баварія) на розробках сланцевих пісковиків, що становлять відкладення мілководної затоки юрського моря (вік близько 150 млн. років). Їх назвали Archaeopteryx lithographicaі Archaeornis simetisi. У 1956 р. там же (в радіусі до 300 ж, але на іншій глибині) знайдено значно гіршої безпеки відбиток ще однієї особини. Частина дослідників розглядає ці знахідки як залишки трьох різних видів (два види роду Archaeopteryx і один роду Archaeornis), інші вважають представниками одного виду Archaeopteryx lithographicaпояснюючи відмінності між особинами як індивідуальні, вікові та статеві. Питання це потребує додаткового вивчення, але й зараз цілком очевидна дуже велика близькість всіх цих особин. Всі вони мають добре розвинене оперення крил, хвоста та тіла, деякі пропорції черепа ближче до пташиних, ніж до рептильних, пташині риси несуть пояс передніх кінцівок та таз, задні кінцівки. Типово рептильні риси: немає рогової дзьоба, є слабкі зуби, кисть несе першорядні махові, але пряжка ще не утворилася і є три добре розвинені рухомі пальці з великими кігтями, утворення складного крижів тільки почалося (хребці ще не зростаються, рухливі кістки з'єднуються з хребців), стегно причленовується під тупим кутом (направлено кілька вбік), проксимальні тарзальні кістки ще не приросли до кінця великої гомілкової кістки, добре розвинена мала гомілкова кістка, не завершено утворення цівки і т. п. За зовнішнім виглядом, рядом внутрішніх морфологічних ознак і Наскільки це можна припускати зараз, спосіб життя архептерикс (і археорніс) - стародавні, примітивні, але птахи.
Підклас справжніх, або віялохвостих, птахів Neornithes, мабуть, - прямі нащадки якихось ящерохвостых птахів Archaeornithes. Деякі дослідники (наприклад, Гадов) вважають за прямих предків птахів археоптериксу і археорнісу, інші (їх, мабуть, більшість) розглядають групу археоптериксу як примітивну, але спеціалізовану бічну та сліпу гілку, вважаючи, що предками сучасних птахів мають бути інші, поки що не виявлені ящерохвості птахи (у разі їх виявлення вони, ймовірно, утворюють інший загін цього підкласу).
Походження та взаємини сучасних груп (загонів) птахів залишаються, як говорилося, за відсутністю достатніх палеонтологічних матеріалів, зазвичай, дуже гіпотетичними. Найбільшу увагу приділив цьому питанню М. Фюрбрінгер, помітно менший - Г. Гадов. Не внесли багато нового і безперечного в цю проблему і сучасні автори, які тією чи іншою мірою стосувалися філогенії птахів. Тому тут викладено уявлення про взаємини окремих груп птахів переважно по Фюрбрінгера, але в ряді випадків, де це потрібно, внесено деякі зміни (у окремих випадках ці зміни спеціально не обумовлюються).
Викопні залишки віялохвостих птахів виявлені тільки в крейдяному періоді (їх вік приблизно 80 - 90 млн. років), тобто вони на кілька десятків мільйонів років молодші за археоптерикси. За декільком поганим збереженням залишкам описані гесперорніси - дуже великі (довжиною понад 100 см) птиці, зовні, мабуть, дещо схожі на гагар або поганок, які зовсім не літали (грудина без кіля, від передньої кінцівки збереглося тільки маленьке тоненьке плече), але, ймовірно , вони добре плавали і пірнали: щелепи їх несли зуби. У крейдяних відкладеннях знайдено залишки інших зубастих птахів - іхтіорнісів. Вони мали добре розвинений кіль грудини, типове пташине крило і, мабуть, мали активний політ. За розмірами вони були приблизно з голуба. Споріднені стосунки зубастих крейдяних птахів з іншими птахами дуже незрозумілі. Можливо, частина цих залишків відноситься до крейдових динозаврів, а не птахів, але частина, безперечно, зубастих птахів. Фюрбрінгер зближував гесперорнісів з гагарами та поганками, а їх тіорнісів – з чайками, але при цьому підкреслював їхню дуже давню відокремленість.
Гадов гесперорнісів виділяв у надзагін, показуючи цим їхнє відокремлене становище, а їхтіорніс як самостійний загін (теж відокремлений) починали перелік загонів кілегрудних птахів. Наголошуючи на неясності походження (та й обсяг) цих груп, Уетмор (1960) виділяє їх у самостійні надзагони.
З крейдових відкладень описано ще кілька сімейств птахів (за уламками щелеп та трубчастих кісток), але їх становище та зовнішній вигляд дуже незрозумілі: можливо, хоча б частина цих знахідок – залишки рептилій. Зустрічені у відкладеннях третинного періоду кайнозойської ери (вік менше 55 млн. років) залишки птахів із більшою чи меншою ймовірністю можуть бути віднесені до сучасних загонів. У всякому разі вони належать до типових птахів, близьких до тих чи інших груп сучасних птахів. Судячи з цих матеріалів, можна припускати, що інтенсивна адаптивна радіація птахів та становлення більшості сучасних загонів проходило наприкінці крейдяного періоду мезозойської ери - в третинний період кайнозойської ери, тобто в період приблизно 70 - 40 млн років тому.

Гадів усіх нині живих і копалин великих нелітаючих страусоподібних птахів виділяв у загін безкільових, підкреслюючи цим їхню подібність і філогенетичне спорідненість (тобто. монофілію), і навіть їх певну відособленість з інших груп птахів. Серед кільових птахів найбільш близькі до страусоподібного тинаму (скритохвости), курячі, журавлеподібні та близькі до них групи. Фюрбрінгер і більшість сучасних авторів (Уетмор, Штреземанн та ін.) вважають страусоподібних птахів збірною поліфілетичною групою, що розбивається на ряд самостійних загонів, що починають систему, що передбачає їхню певну примітивність та ізольованість. Їхні спільні подібні риси - результат конвергенції: великі розміри, втрата здатності до польоту, пристосування до швидкого бігу і т.д.
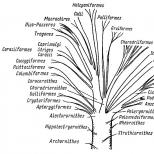
По Гадову, еволюція інших птахів у двох напрямах (дві гілки, див. вище), кожен із яких у своє чергу поділяється на 2 групи родинних загонів. Поганко-гагарообразные (ихтиорнисы, поганки, гагары, пингвины и трубконосые) родственны группе Pelargomorphae (аистообразные, гусеобразные, хищники), а близкая к страусоподобным группа курообразных (тинаму, куриные, журавлеобразные, куликообразные) дала начало ракшеобразным (кукушки, ракши, воробьиные) .
Еволюційні уявлення Фюрбрінгера в загальних рисах близькі до цієї схеми Гадова (про інші погляди на страусоподібних говорилося вище), але він викладає свої уявлення з більшим ступенем деталізації. Великий стовбур птахів - загін Pelargornithes - ділиться на ряд гілок: гусеподібних, поганкоподібних (включаючи поганок, гагар, гесперорнісів); від заснування цих гілок відходить ще одна потужна гілка, яка, поділяючись, дає початок веслоногим, денним хижакам та групам підряду лелеподібних (рис. 14).
Загін паламедей Palamedeiformes займає проміжне положення між нанду і загоном Pelargornithes. Проміжні загони пінгвінів Aptenodytiformes і трубконосих Procellariiformes досить відокремлені; можливо, вони мають давні слабкі зв'язки один з одним і, ще більш віддалені, з загонами Pelargornithes і куликоподібних Charadriornithes. Іхтіорніс Ichthyornithiformes - теж проміжний загін, але з дещо більш вираженими давніми зв'язками з куликоподібними. Загін куликоподібних Charadriornithes дуже давніми зв'язками мабуть може бути наближений до загону ракоподібних. Основні розгалуження стовбура куликоподібних - підзагін Laro-Limicolae - кулики, чайки та чистики; до них досить близькі зобаті бігунки і дрохви (перехідною групою між кулями і дрохвами служать авдотки). Від основи кулікаподібних відокремилися ще дві великі гілки: журавлеподібні Gruiformes і пастушкові Ralliformes. Журавлеподібні поділяються на кілька гілок: власне журавлиних (журавлі, араміди, трубачі, каріами) і займають дещо відокремлене становище сонячних чапель та кагу. Пастушкові поділяються на споріднені гілки: пастушків, лапчатоногих і дещо окремо триперсток і мадагаскарських пастушків.
Один з древніх стовбурів родоводу птахів - загін куроподібних альекторорнітів. Найбільш примітивна гілка - підряд безкрилих, або ківі Apterygiformes, що включає ківі і моа; обидві групи розійшлися майже одразу після свого відокремлення. Загальним походженням з попередньою гілкою, мабуть, пов'язані тинаму (або приховані хвости) Crypturiformes, у свою чергу через предків близькі до підряду курячих Galliformes (практично в сучасному обсязі). Загін куроподібних дуже давніми родинними зв'язками, ймовірно, більш-менш близький до пастушкових Ralliformes, а через них і до всього загону куликоподібних.
Проміжні загони голубоподібних Columbiformes (включаючи брижів і голубів) і папуг Psittaciformes займають у системі відокремлене положення. Ймовірно, через дуже давні стосунки вони якось пов'язані з великим загоном ракоподібних. За уявленнями Гадова, голуби і рябки в ранзі підзагонів входять у загін кулькоподібних, а папуги - теж як підзагін - у загін зозулеподібних. По суті справи подібної точки зору дотримується і Уетмор, що ставить загін голубоподібних відразу ж за загоном куликоподібних, а загін папуг - поряд з загоном зозулів.
Великий і різноманітний загін Coracornithes своїми витоками пов'язаний з предками загону куликоподібних. Основна і найбільш потужна гілка - підряд дятлів і горобців Pico - Passeriformes, яка потім поділяється на окремі групи власне горобиних і дятлових. Від заснування цієї гілки відокремилися Macrochires (стрижі, колібрі) та птахи миші Colii. Ще раніше від цієї галузі відокремлюються трогони Тгоgones. Від самої основи гілки дятлово-горобиних птахів відокремлюються і три інших підряди ракоподібних. Підзагін зозулеподібних Coccygiformes відокремлює гілку якамар Galbulae (якамари і пухівки, а по Уетмору - це підзагін загону дятлів), а потім поділяється на зозулі Cuculidae і бананоїдів Musophagidae. Подотряд ракшеподібних Со- raciiformes поділяється на власне ракш Coraciae і на другу гілку, що незабаром поділяється на сов Striges і козодоїв Caprimulgi. Підзагін зимородкових Halcyoniformes (він, мабуть, ближче до Pico - Passeriformes, ніж до інших підзагонів), відокремлюючи на самому початку бічну гілочку тоді Todi (крім тоді Фюрбрінгер сюди включає і момотів), поділяється на 3 родинні групи: зимородкових Halcyones (Al ), удодів Bucero tes (удоди, птахи-носороги) і щурок Meropes.
Зіставлення філогенетичних уявлень Фюрбрінгера з поглядами Гадова і Уетмора (черговість і порядок перерахування загонів) показує значний їхній збіг; різкі відмінності у положенні та взаємовідносинах окремих груп зазначені вище.

В останні роки спробу графічного зображення філогенетичних взаємин між окремими групами птахів зробили німецькі орнітологи Берндт і Майзе в капітальному тритомному зведенні "Природна історія птахів" (Bernd R., Meise W. Naturgeschichte der Vogel, Kosmos, Stuttgart, 1, 9). , Стор 668 - 673). Підклас древніх ящерохвостых птахів Palaeornithes дав початок підкласу нових віялохвостих птахів Neornithes, який включає 24 загони, які підрозділяються авторами зазвичай відразу на сімейства; Таксономічні категорії рангу підзагону та надродини використовуються, як правило, тільки в загоні гороб'їних. Взаємини між загонами птахів, по Берндту та Майзі, виглядають так:
Від древніх птахів еволюція йшла по двох напрямках (гілках): гілки (надзагін) наземних і водних птахів Георнітх і гілки (надзагін) деревних птахів Dendrornithes. Від самого початку гілки наземних і водних птахів (мал. 15) загальним стовбуром відокремилися, пізніше розділившись, тинаму Crypturi і безкілеві птахи Ratitae, включаючи всіх страусоподібних, що нині живуть, у тому числі і ківі, і копалин моа і епіорнісів. Далі відокремлюються курячі птахи Galli (в повному обсязі загону Уетмора) і на цьому ж рівні, але самостійною гілкою - гагароподібні Pygopodes (що включають викопні крейдяні зубасті птахи гесперорнісів, гагар і поганок). Всі інші загони цієї гілки (надзагону) мають загальне походження - являють собою як би мутовку стовбурів, що відходять від однієї ділянки. Відносно тісно пов'язані один з одним і дещо протиставляються іншим групам загони журавлеподібних (пастушки, мада-гаскарські пастушки, сонячні чаплі, лапчатоноги, кагу, справжні журавлі, арами, трубачі, дрохви, каріами, трифорки і скопа; болотно- і чайкоподібних птахів Limicolae - Lari (якани, різні групи власне куликів, чайки чистики та - з копалин - зубасті крейдяні птахи - іхтіорніси). Далі від загальної основи мутовки (тобто маючи спільних предків) послідовно відокремлюються загони гусеподібних Anseres (паламедеї і власне гусеподібні), фламінго Phoenicopteri, гомілі Gressores, денні хижаки Accipitres і веслоногі Steganopodes. Від форм, близьких до предків веслоногих, відокремлюється гілка, яка потім поділяється на загони трубконосих Tubinares та пінгвінів Sphenisci.
Розвиток другої гілки (надзагону) деревних птахів Dendrornithes починається від якихось предкових форм, мабуть, більш давніх, ніж предки першої гілки (першого надзагону). Спочатку загальним стовбуром відокремлюються 3 загони: зозулеподібних Cuculi, голубів Columbae (голубки, рябки та дронти) та папуг Psittaci. Наступна відокремлена гілка поділяється на 2 відгалуження, кожне з яких утворює по 2 загони. Одне з них дає початок совам Striges та козодоям Caprimulgi, інше – трогонам Trogones та ракоподібним Coracii. Далі від загального стовбура також загальною гілкою відокремлюються загони птахів-мишей Colii та стрижеподібних Macrochires (стрижі та колібрі), самостійною гілкою відходить загін дятлів Pici (в об'ємі Уетмора), а основний стовбур розгалужується на численні підрозділи загону горобиних Pass. Крім цього, автори дають гіпотетичні схеми, в яких намагаються подати філогенетичні взаємини в межах кількох окремих загонів (денних хижаків, веслоногих, гагароподібних та гороб'ячих).
Філогенетичні схеми Берндта та Майзе зрозумілі, наочні та легко запам'ятовуються. Однак вони практично зовсім не аргументовані, не обґрунтовані матеріалами і, мабуть, значно меншою мірою відображають дійсні споріднені (філогенетичні) відносини між окремими групами птахів, ніж відповідні схеми Фюрбрінгера. Теоретично малоймовірно, що вже на початкових стадіях еволюції класу чітко визначилася наступна біотопічна спеціалізація птахів, що виражається у викладеній схемі поділу підкласу "нових птахів" тільки на 2 відокремлені надзагони та в майже лінійному, послідовному відокремленні загонів. Оцінюючи за сучасними формами властивий птахам характер еволюції, що виражається у широкій адаптивній радіації в межах окремих груп (про що коротко згадувалося вище), здається більш логічним припускати, що подібна широка адаптивна радіація була характерна і для початкових фаз еволюції класу. Якщо це так, то можна говорити, що філогенетичні схеми Фюрбрінгера хоча б зовні справді відбивають переважання такого типу еволюції.
Зараз ще рано говорити про систему класу, що цілком склалася. Однак системи Фюрбрінгера, Гадова та Уетмора, мабуть, можуть розглядатися як хороша основа для побудови природної системи. Їхні матеріали дають і найзагальніше уявлення про філогенетичні взаємини більшості груп. Для покращення та уточнення системи класу птахів необхідні подальші різноманітні дослідження на всіх таксономічних рівнях (від підвидових угруповань до загонів та надзагонів): морфологічні, екологічні, етологічні, генетичні. Можливо, у майбутньому цінні матеріали дасть застосування біохімічних та фізіологічних показників; використання сучасних методик поки що істотного внеску в таксономію птахів не внесло. Хочеться сподіватися, що збільшиться і кількість палеонтологічних матеріалів, які дозволять більш обґрунтовано обговорювати еволюцію птахів загалом та окремих груп (загонів, родин).
Сучасні птахи сильно відрізняються від родинних хребетних. Донедавна їхнє походження було однією з великих таємниць біології. Пір'я, беззубі дзьоби, порожнисті та глибокі грудні кістки є лише частиною особливих ознак, яких немає в інших тварин. Спостереження за птахами не пояснює, як у них з'явилося пір'я та здатність до польоту. За останні десятиліття з'явилися нові відкриття та нові методи дослідження, які дозволили палеонтологам зробити кілька відкриттів. Знайдені в районі Китаю, Південної Америки скам'янілості, а також зразки, що зберігаються в музеях, які можна вивчати за допомогою нових технологій, стали причиною виникнення чергової хвилі інтересу до вивчення історії еволюції птахів. Завдяки цьому, раніше з'явилася теорія їхнього походження від дрібних мешканців Землі наприкінці юрського періоду.
Початок дослідження виникнення птахів
Вчені почали міркувати про еволюцію птахів, починаючи з того часу, як Чарльз Дарвін виклав свою теорію еволюції у роботі «Про походження видів». У 1861 році, через рік після публікації трактату Дарвіна, древнє пташине перо було знайдено в баварських вапнякових відкладах, вік яких становив близько 150 млн років. Наступного року було знайдено скелет тварини, яка мала пташині крила і пір'я, дуже довгий кістлявий хвіст і щелепу із зубами. Він був знайдений у тому ж регіоні. Скам'янілий кістяк був названий археоптериксом. Він став першою знайденою найдавнішою твариною з оперенням. Скелетна анатомія археоптерикса давала чіткі докази, що предками птахів були динозаври, але 1861 року вчені ще змогли встановити цей зв'язок. Тоді й почалося вивчення еволюції птахів та полювання за предками сучасних видів пернатих.
Анатомічна будова археоптериксу
Скам'янілість було виявлено на початку 1860 років. Довгий час знаменита знахідка з пізньої юри була унікальною. Вона єдина могла дати якусь інформацію у тому, як міг статися еволюційний перехід від рептилій до птахів, оскільки у ній поєднувалися ознаки як птахи, і рептилії. Будівля археоптерикса мала багато спільного з будовою птахів. Наприклад, пір'я вздовж передніх лап, що стали крилами. Але, на відміну від сучасних видів, також були зуби і костистий хвіст.
Перші теорії про тварин юрського періоду
Варто відзначити, що багато кісток тварини, у тому числі передніх лап, плечового пояса, тазу і ніг, були виразними, не зрослими, як у його нащадків. Через кілька років Томас Генрі Хакслі став першим ученим, хто знайшов зв'язок між будовою птахів і динозаврами. Він порівнював задні кінцівки гігантського динозавра із зображенням страуса і відзначив 35 ознак, що доводять, що вони пов'язані одна з одною. Хакслі представив свої результати Геологічному суспільству. Але в 1870 році в Лондоні палеонтолог Гаррі Говє Сілі вирішив оскаржити гіпотезу походження птахів та їх спорідненість із динозаврами. Силі припустив, що задні кінцівки страуса і динозавра можуть виглядати схожими лише тому, що ці тварини були великими і двоногими, а свої задні кінцівки використовували у подібних умовах. Крім того, динозаври були навіть більшими, ніж страуси, і ніхто з них не міг літати. У вченого постало питання, як тоді можуть літати птахи, якщо вони еволюціонували від динозавра.

Теорія Герхарда Хайльмана
Таємниця знову зацікавила вчених приблизно через півстоліття. У 1916 році Герхард Хайльман, медичний лікар, який цікавиться палеонтологією, опублікував датською мовою блискучу книгу про історію походження птахів, яка в 1926 була перекладена англійською під назвою "Походження птахів". Хайльман показав, що пернаті були анатомічно більш схожі на динозаврів-тероподів, ніж будь-яку іншу викопну групу. Але існувала одна неминуча невідповідність: у тероподів явно були відсутні ключиці, ті, що зливаються у вилочку у птахів.
Недоліки теорії Герхарда Хайльмана
Через той факт, що інші рептилії мали ключиці, Хайльман припустив, що тероподи, зокрема тріасові псевдозухії, з якоїсь причини втратили їх. Для нього ця втрата означала, що птахи не могли розвинутись від цього виду динозаврів. Тому він був переконаний, як пізніше з'ясувалося, помилково, що функція ключиць, втрачена під час еволюції, не може бути відновлена. Птахи, стверджував він, мали розвинутися з більш архаїчної рептильної групи, яка мала ключиці.

Як раніше Силі, Хайльман дійшов висновку, що подібність між птахами та динозаврами має просто відображати той факт, що обидві групи були двоногими. Його висновки вплинули на палеонтологічні дослідження і протягом тривалого часу були пріоритетними, хоча нова інформація і спростувала деякі факти. Але досі деякі вчені дотримуються саме її. Два окремі дослідження показали, що тероподи справді мали ключиці. У 1924 році було опубліковано анатомічний малюнок незвичайного тероподу з папужним оперенням. Він був названий овіраптором, і його існування спростовувало теорію Хайльмана.

Нові докази
У 1936 році Чарльз Камп з Каліфорнійського університету в Берклі знайшов останки невеликого тероподію раннього юрського періоду і хотів поставити крапку в історії з ключицями. Але доказ хибності теорії Хайльмана мало ким визнано. Нещодавні дослідження знайшли ключиці у багатьох тероподів. Виходячи з будови птахів в даний час, а також внаслідок низки досліджень, археоптерикс був названий проміжною ланкою в історії еволюції пернатих. Але лишалося питання: між якими саме рептиліями? Подальші копалини птахи знайдені в шарах нижче за ранню крейду. Більшість із них було виявлено з початку 1990 років, головним чином у Китаї. Тим часом відомо понад тридцять видів птахів з крейдяного періоду. Крім того, було виявлено велику кількість маленьких динозаврів, що пересуваються на двох ногах із характерними для древніх птахів рисами. Таким чином, розрив між динозаврами та птахами завдяки вивченню скам'янілостей став значно меншим.
Теорія Острома
Нарешті, через століття після спірної презентації Хакслі для Геологічного суспільства Лондона, Джон Х. Остром з Єльського університету відродив ідею, що птахи були пов'язані з тероподами. Він прямо припустив, що пернаті є їх прямими нащадками. Наприкінці 1960-х років Остром описав скелетну анатомію хижого теропода дейноніха, який був розміром з людського підлітка і жив на Землі близько 115 мільйонів років тому, в епоху ранньої крейди. У подальших своїх опублікованих роботах Остром продовжив визначення анатомії птахів та групи особливостей, які вони, включаючи археоптерикс, ділили з дейноніхом та іншими тероподами, але не з іншими рептиліями.
На основі цих даних він дійшов висновку, що птахи походять безпосередньо від дрібних динозаврів-тероподів. Після висування теорії Остром продовжував збирати свої докази походження пернатих від тероподів. Він використав новий метод розшифрування відносин між організмами, застосовуючи його в музеях природної історії у Нью-Йорку, Парижі та інших місцях. Цей метод отримав назву кладистики чи філогенетичної систематики. Оскільки він став стандартом для порівняльної біології, його використання суворо доводило висновки Острома.

Дослідження 1970-х років
У 1970-х роках вчені продовжували цікавитись деякими загальними унікальними особливостями археоптерикса, які ріднять його з деякими тваринами юрського періоду. Серед них опинилися ті найменші м'ясоїдні динозаври, що пересуваються на двох ногах, які отримали назву тероподів. Уважно вивчивши ці риси, вчені знову висунули теорію, що, можливо, вони були предками птахів. Провідні палеонтологи збудували еволюційні дерева, і після цього виявилися ще більш переконаними у своїй правоті.
Що таке кладограма
Ці дерева, інакше відомі як скарбниця, є сучасним золотим стандартом в аналізі еволюційних відносин між тваринами. Сам метод отримав назву кладистика. Практикуючі кладистики визначають еволюційну історію групи тварин, вивчаючи різні види ознак. У процесі еволюції в якоїсь тварини може з'явитися нова, генетично певна риса, яка буде передана його нащадкам. Отже, палеонтологи можуть зробити висновок, що дві групи, однозначно поділяють набір таких нових або отриманих рис, більш тісно пов'язані один з одним, ніж з тваринами, які цих рис позбавлені. Вузли або точки розгалуження на скарбниці вказують на появу лінії з новим набором похідних ознак. В результаті птахи здалися вченим просто гілкою на дереві життя динозаврів. У роботах Дж. Острома в середині 1970-х років тероподи набули статусу найімовірнішої групи для предків птахів. Тому сьогодні ідея, що птахи є пернатими динозаврами, практично не заперечується серед еволюційних теоретиків.
Особливості методу кладистики
Традиційні методи угруповання організмів за принципом подібності та відмінності можуть виключити вид із групи тільки тому, що він має риси, які не були знайдені в інших членів. На противагу цьому групи кладистики були засновані виключно на певних групах загальних рис, які особливо інформативні. Цей метод починається з дарвінівської заповіді про те, що еволюція продовжується, коли з'являється нова спадкова риса в деяких організмах, яка і передається генетично його нащадкам. Припис вказує, що дві групи тварин, що поділяють такі нові риси, тісніше пов'язані один з одним, ніж ті, що поділяють тільки оригінальні, але не похідні, передані у спадок. Визначаючи загальні похідні риси, практики-кладистики можуть визначати відносини серед організмів, що вивчаються. Результати таких аналізів, які зазвичай досліджуються, можуть бути представлені у вигляді кладограми. Деревоподібна діаграма, що зображує порядок, у якому з'являлися нові характеристики та нові істоти, відбивають той порядок, у якому відбувалася еволюція.
Причина появи пір'я у птахів
У міру еволюції птахів та їх віддалення від динозаврів, названих тероподами, безліч їх особливостей змінювалися та вдосконалювалися. Якщо придивитися уважніше, стає цілком очевидно, що набір цих характерних ознак довго розвивався і служив для виконання певної функції. Наприклад, поява оперення у маленького тероподу було пов'язане з необхідністю ізоляції певних ділянок шкіри і спочатку більше нагадували волосся. На першому пташиному пір'ї, можливо, були різні кольорові візерунки. Існує версія, що вони були призначені для камуфляжу, розпізнавання свого виду та інших функцій.
Порівняння анатомічних структур птахів та динозаврів
Порівняльна анатомія птахів та тероподів допомогла пов'язати їх один з одним. Вона також виявила деякі способи зміни цих характеристик, оскільки динозаври стали ближчими до птахів, а птахи виявилися сучаснішими. Наприклад, в ділянці таза лобкова кістка, спочатку спрямована вперед, пізніше зміщується вертикально або назад. У передніх лапах відносні пропорції кісток залишалися досить незмінними у ранніх птахів, але зап'ястя змінилося. У деяких видів кістка у зап'ясті набула форми півмісяця. У результаті її форма сприяла можливості злету. Широка вилочка бумерангообразная у перших оперних динозаврів стала тонше і утворила глибшу дугу, коли ця особливість стала необхідна для польоту.
Загальні ознаки динозаврів та птахів
Дослідження Готьє, проведені останнім часом, демонструють, що багато функцій, які традиційно вважаються такими, що належать тільки птахам, фактично з'явилися до них, у їхніх предків — тероподів. Багато з цих властивостей допомогли своїм первісним власникам вижити. Ці ж риси та деякі інші були зрештою використані чи перетворені для польоту та життя на деревах. Пташині характеристики тероподів, які еволюціонували до птахів, не виявлялися одразу, а деякі були до того, як з'явилися самі тероподи. Вони були ще у динозаврів, які існували раніше. Наприклад, безпосередній предок тероподів був двоногим і пересувався як птах. Він був маленьким і м'ясоїдним. У нього були передні лапи, як і ранні птахи. Крім того, другий, а не третій палець, як у інших рептилій, був довшим. У предків динозаврів гомілковостопний суглоб став шарнірним, а кістки стопи подовженими. Багато змін у будові ніг, як вважають, вплинули збільшення довжини кроку і швидкість бігу. Ця властивість одного прекрасного дня допоможе пташиним тероподам злетіти.

Найбільший у світі літаючий птах і найменший
Гігантські птахи мешкали на Землі понад 6 млн. років тому. Найбільшим у світі літаючим птахом був аргентавіс. Він мешкав в Аргентині і належав до сімейства соколиних. Дорослий птах досягав 2 метрів у висоту, а його череп був приблизно півметра в довжину. Величезні крила у розмаху були як триповерховий будинок. Найменший птах існує і зараз. Це широко відомий вид - колібрі-бджілка. Довжина його тіла зазвичай не перевищує 6 см. Місце проживання колібрі - степові райони бразильської провінції Мінас-Жерайс. Найменший птах харчується нектаром квітів і дуже швидко махає крилами, як бджола, тому й отримав таку назву.
Риси подібності між птахами та плазунами
Легко переконатися у подібності між класами птахів та плазунів. У тих та інших шкіра майже позбавлена залоз, але захищена роговими лусочками – у плазунів та пір'ям – у птахів. Зазначимо, що птахи лусочки розвинені на неоперених частинах шкіри (цівках). Пір'я птахів також є роговими утвореннями, що розвинулася з луски. Обидва класи відносяться до яйцекладних, і яйця влаштовані схожим чином: шкаралупа, жовток та білок. Зародки птахів і плазунів схожі на вигляд.
Предки птахів – стародавні рептилії
У пошуках безпосередніх предків-рептилій були обрані дрібні примітивні плазуни псевдозухіїякі жили приблизно 200 млн. років тому (тріасовий період). У пошуках їжі деякі з цих істот пристосовувалися до лазіння по деревах і стрибках з гілки на гілку. У ході еволюції цей спосіб виявився перспективним і допоміг первісним птахам уникати конкуренції серед родинних видів та рятуватися від хижаків. У міру подовження лусок, вони перетворювалися на пір'я, що допомогло древнім предкам птахів набути здатність до плануючого, та був і до активного, тобто. махаючого, польоту, яку мають більшість сучасних птахів.
Найдавніший птах
Protoavia (1984 р.) було знайдено у містечку Пост, Техас, США, її гаданий вік 225 000 000 років.
Перші птахи з'явилися у мезозойську еру
Розвиток Землі поділено п'ять проміжків часу, які називаються ерами. Перші дві ери, археозою та протерозою, тривали 4 мільярди років, тобто майже 80% усієї земної історії. Під час археозою сталося утворення Землі, виникла вода та кисень. Близько 3,5 мільярдів років тому з'явилися перші крихітні бактерії та водорості. В епоху протерозою близько 700 років тому в морі з'явилися перші тварини. Це були примітивні безхребетні істоти, наприклад хробаки та медузи. Палеозойська ера почалася 590 мільйонів років тому і тривала 342 мільйони років. Тоді землю покривали болота. Під час палеозою з'явилися великі рослини, риби та земноводні. Мезозойська ера почалася 248 мільйонів років тому і тривала 183 мільйони років. У цей час населяли Землю величезні ящери динозаври. З'явилися також перші ссавці та птахи. Кайнозойська ера почалася 65 мільйонів років тому і продовжується досі. У цю пору виникли рослини та тварини, які оточують нас сьогодні.
Сталися від целурозаврів

Ті, що жили в кінці тріасу і в юрському періоді, невеликі хижі динозаври з групи целурозаврівбули двоногими з довгими хвостами та дрібними передніми кінцівками хапального типу. Їм не було потреби лазити по деревах і планувати з гілки на гілку. Активний політ древніх птахів міг виникнути на основі рухів передніх кінцівок, що плескали, що допомагало збивати летких комах, для чого, до речі, хижакам доводилося високо підстрибувати. Целурозаври пережили масове вимирання динозаврів наприкінці мезозойської епохи.
Найперші птахи-динозаври
У мезозойську епоху, тобто 150 млн років тому, предки птахів становили основну групу наземних тварин біля Аргентини. Їх називають тероподи (Argentavis magnificens),звіроногі ящіри, і вони вже вміли літати. Пересувалися тераподи на двох ногах, передні лапи перетворилися на короткі хапальні кінцівки. Спиратися на них було вже не можна, зате боротися зі здобиччю було зручно. Потужні щелепи тероподів були густо усіяні зубами і нагадували полотно пили. На місці зубів, що зіткнулися, виростали нові, тому, навіть зістарившись, ящери все з тим же запалом могли мучити видобуток. (Також оновлюються зуби і в акул.) У процесі еволюції у деяких тероподів з'явився роговий дзьоб. Аналізуючи анатомічні особливості тероподів, вважають, що птахи походять саме від цих тварин.

Викопні залишки, знайдені в Аргентині в 1979 р., свідчать про те, що у цієї величезної, схожої на грифа птиці розмах крил перевищував 6 м, зростання досягало 7,6 м, а вага - 80 кг
Звіроподібний хижий ящір орнітолест, що мав довжину тіла 2,5 м, дає уявлення про давніх птахів
розмах крил вони мали 7,5 м; вони мешкали в Європі, Африці, Північній та Південній Америці і були м'ясоїдними (їли рибою та водними безхребетними).
Сучасний птах, який жив 120 млн років тому
 Вивчивши скам'янілі останки невеликого птаха, знайдені в провінції Ляонін на півночі Китаю, вчені дійшли висновку, що "конфуціузорніс санктус" - так охрестили стародавнього птаха - жив 120 мільйонів років тому. Судячи з будови дзьоба, конфуціузорніс нагадував сучасних нам птахів: зуби вже були відсутні, натомість виник роговий чохол.
Вивчивши скам'янілі останки невеликого птаха, знайдені в провінції Ляонін на півночі Китаю, вчені дійшли висновку, що "конфуціузорніс санктус" - так охрестили стародавнього птаха - жив 120 мільйонів років тому. Судячи з будови дзьоба, конфуціузорніс нагадував сучасних нам птахів: зуби вже були відсутні, натомість виник роговий чохол.
Поява махаючого польоту
 У юрському періоді птахи набули здатності до активного польоту. Завдяки помахам передніх кінцівок вони змогли подолати дію земного тяжіння та отримали масу переваг перед своїми наземними, лазаючими та плануючими конкурентами. Політ дозволив їм ловити у повітрі комах, ефективно уникати хижаків і вибирати найбільш сприятливі життя середовища. Його розвиток супроводжувався укороченням довгого хвоста із заміною його віялом з довгого пір'я, добре пристосованим для рулювання та гальмування. Більшість анатомічних перетворень, необхідні активного польоту, завершилися до кінця раннього крейди (близько 100 млн. років тому вони), тобто. задовго до вимирання динозаврів.
У юрському періоді птахи набули здатності до активного польоту. Завдяки помахам передніх кінцівок вони змогли подолати дію земного тяжіння та отримали масу переваг перед своїми наземними, лазаючими та плануючими конкурентами. Політ дозволив їм ловити у повітрі комах, ефективно уникати хижаків і вибирати найбільш сприятливі життя середовища. Його розвиток супроводжувався укороченням довгого хвоста із заміною його віялом з довгого пір'я, добре пристосованим для рулювання та гальмування. Більшість анатомічних перетворень, необхідні активного польоту, завершилися до кінця раннього крейди (близько 100 млн. років тому вони), тобто. задовго до вимирання динозаврів.
Про пряму спорідненість ящерів і птахів не може бути й мови

Встановили американські вчені із університету штату Північна Кароліна. Вони порівняли зміни кінцівок тероподів з еволюцією крил курей, страусів та бакланів. У процесі еволюції і ті, й інші зберегли лише по три пальці з первісних п'яти. Однак американські біологи з'ясували, що у птахів відсутні обидва зовнішні пальці, тобто перший і п'ятий. Ящери втратили четвертий і п'ятий пальці.
Довгий птах
Скелет ящера "уненлагіа комахуенсіс", "довгого птаха", знайдений у травні 1996 року в Аргентині, заповнює лакуну, що розділяє стародавні рептилії тероподів і першу птицю арехеоптерикса.
Відмінності між птахами та плазунами
Про вищому розвитку птахів свідчать збільшений головний мозок (зокрема, великі розміри великих півкуль та мозочка у птахів), досконалість дихальної та кровоносної систем – подвійне дихання та відділення артеріальної крові від венозної, а також постійна температура тіла. Всі ці вдосконалення організації птахів відсутні у плазунів.
Найдавніший птах літав
Суперечка, чи міг археоптерикс літати, триває з 1861 р., коли було знайдено першу копалину, і досі. Відповідь було знайдено лише недавно. Скам'янілий мозок істоти помістили в рентгенівську установку, що дозволяє отримувати тонкі зрізи об'єкта. Ці зрізи об'єднувалися в комп'ютері в тривимірну модель. Виявилося, що за своєю анатомією мозок археоптерикса набагато ближчий до мозку сучасних літаючих птахів, ніж до мозку динозаврів, як передбачали палеонтологи раніше. Аналіз показав, зокрема, напівкруглі канали у внутрішньому вусі, що використовуються для балансу, та збільшені частки мозку, що відповідають за зір – особливості, що впливають на ефективність польоту. "Літаючий" мозок розвивався одночасно з крилами, а сама здатність літати виробилася в процесі еволюції набагато швидше, ніж вчені вважали раніше.
Кондиціонером служили кістки
Під час полювання древні ящероптиці тероподи регулювали температуру свого тіла, щоб уникнути перегріву. Функцію кондиціонера виконували порожнисті кістки.
Археоптерикс – безпосередній прабатько сучасних птахів
 Останки вимерлої птиці, що нагадує із сороку, що жила в другій половині юрського періоду, тобто 100%. 140 млн. років тому були відкриті в Європі. У шарах земної кори вчені виявили скам'янілі кістки скелета невідомої істоти, а поряд відбитки його пір'я. Птах отримав ім'я археоптерикс (Archaeopteryx litographica),що означає «давній птах». Ця невелика пташка мала гострі зуби в осередках, довгий, як у ящірки, хвіст і передні кінцівки з трьома пальцями, що несли гачкуваті пазурі.
Останки вимерлої птиці, що нагадує із сороку, що жила в другій половині юрського періоду, тобто 100%. 140 млн. років тому були відкриті в Європі. У шарах земної кори вчені виявили скам'янілі кістки скелета невідомої істоти, а поряд відбитки його пір'я. Птах отримав ім'я археоптерикс (Archaeopteryx litographica),що означає «давній птах». Ця невелика пташка мала гострі зуби в осередках, довгий, як у ящірки, хвіст і передні кінцівки з трьома пальцями, що несли гачкуваті пазурі.
Археоптерикс нагадував плазуне
Форма черепа археоптерикса із зубами в обох щелепах і дуже довгий хвіст із 20 хребцями нагадували плазуне. Більшістю ознак археоптерикс скидався більше на плазуна, ніж на птаха, якщо не брати до уваги справжнє пір'я на передніх кінцівках і хвості.
Як літали археоптерикси
Все тіло цієї істоти, крім голови, було вкрите пір'ям, а передні кінцівки мали всі основні особливості пташиних крил з маховим пір'ям. Тільки пальці крила були довшими, ніж у сучасних птахів, і мали пазурі. На ногах було по чотири пальці: перший палець звернений назад, решта – вперед, що допомагало добре обхоплювати пальцями гілки. Рульове пір'я прикріплювалося попарно на кожному хребці довгого хвоста, а не так, як у сучасних птахів, - широким віялом на копчиковій кістці. Особливості арехеоптерикса показують, що він був здатний до польоту, що махає, але тільки на дуже короткі відстані.
Тварина розміром із ворону
 Рахонавіс (Rahonavis)- це тварина розміром із ворону, яка жила близько 80 мільйонів років тому, належить до тієї ж групи динозаврів, що й велоцираптор. Щоправда, істота має дуже багато спільного з птахами. У рахонавісу був серповидний кіготь, що висувається, на середньому пальці ніг, пір'яний покрив і довгий пазуристий хвіст подібно до археоптерикса.
Рахонавіс (Rahonavis)- це тварина розміром із ворону, яка жила близько 80 мільйонів років тому, належить до тієї ж групи динозаврів, що й велоцираптор. Щоправда, істота має дуже багато спільного з птахами. У рахонавісу був серповидний кіготь, що висувається, на середньому пальці ніг, пір'яний покрив і довгий пазуристий хвіст подібно до археоптерикса.
Перші птахи жили у лісі
Перші представники класу виникли і стали опановувати польот, живучи в лісі на гілках дерев, стрибали і лазили по гілках, чіпляючись за них довгими пальцями передніх кінцівок з кігтями. Розправивши крила, вони планували в повітрі зверху вниз похилою площиною, а також пролітали невеликі відстані, змахуючи крилами. Лише пізніше деякі птахи почали пристосовуватися до життя в степах та пустелях, на берегах водойм та інших місцях.
Першоптиця - ще один птахівдінозавр
 Археоптерикс довгий час залишався єдиною відомою науці сполучною ланкою між птахами і рептиліями, проте в 1986 були знайдені залишки ще однієї копалини, яка жила на 75 млн. років раніше і поєднувала в собі ознаки динозаврів і пернатих. Хоча цю тварину й назвали Protoavis (первоптиця),його еволюційне значення викликає суперечки серед учених.
Археоптерикс довгий час залишався єдиною відомою науці сполучною ланкою між птахами і рептиліями, проте в 1986 були знайдені залишки ще однієї копалини, яка жила на 75 млн. років раніше і поєднувала в собі ознаки динозаврів і пернатих. Хоча цю тварину й назвали Protoavis (первоптиця),його еволюційне значення викликає суперечки серед учених.
Багато видів птахів з'явилися в крейдяному періоді
Після археоптерикса в палеонтологічному літописі пернатих спостерігається пропуск тривалістю близько 20 млн. років. Наступні знахідки відносяться до крейдяного періоду, коли з'явилося безліч видів птахів, пристосованих до різних місцеперебування. Серед приблизно двох дюжин крейдових таксонів, відомих по викопних залишках, особливо цікаві два – Ichthyornisі Hesperornis. Обидва були відкриті в Північній Америці, в гірських породах, що сформувалися дома великого внутрішнього моря.
Іхтіорніс – давня чайка
 Іхтіорніс був такої ж величини, як і археоптерикс, його довжина тіла становила близько 50 см, а важив він 5 кг. Зовні він скидався на чайку з добре розвиненими крилами, що вказують на здатність до потужного польоту. Як і в сучасних птахів, зуби у нього були відсутні, але хребці були схожі на риб'ячі, звідки і родова назва, що означає «рибоптиця». Його останки було знайдено у США. Жив їхтіорніс 65-90 тис. років тому.
Іхтіорніс був такої ж величини, як і археоптерикс, його довжина тіла становила близько 50 см, а важив він 5 кг. Зовні він скидався на чайку з добре розвиненими крилами, що вказують на здатність до потужного польоту. Як і в сучасних птахів, зуби у нього були відсутні, але хребці були схожі на риб'ячі, звідки і родова назва, що означає «рибоптиця». Його останки було знайдено у США. Жив їхтіорніс 65-90 тис. років тому.
Гесперорніс – давня гагара
 Гесперорніс («західний птах») був довжиною 1,5-1,8 м (до 2 м) і майже безкрилим. Його вага становила 40 кг. За допомогою величезних ластоподібних ніг, що відходили вбік під прямим кутом на самому кінці тулуба, він, мабуть, плавав і пірнав не гірше за гагар. Він мав зуби «рептильного» зразка, але будова хребців відповідала типовому для сучасних птахів. Останки гесперорнісу знайдені у США. Жив цей птах 70 тисяч років тому.
Гесперорніс («західний птах») був довжиною 1,5-1,8 м (до 2 м) і майже безкрилим. Його вага становила 40 кг. За допомогою величезних ластоподібних ніг, що відходили вбік під прямим кутом на самому кінці тулуба, він, мабуть, плавав і пірнав не гірше за гагар. Він мав зуби «рептильного» зразка, але будова хребців відповідала типовому для сучасних птахів. Останки гесперорнісу знайдені у США. Жив цей птах 70 тисяч років тому.
Сучасні птахи сформувалися 65 млн років тому

З настанням третинного періоду (65 млн. років тому) кількість видів птахів почала швидко збільшуватися. Цим періодом датуються найдавніші копалини пінгвіни, гагари, баклани, качки, яструба, журавлі, сови та деякі співачі.
Величезні птахи, що не літають
Крім предків сучасних видів у третинному періоді з'явилося кілька величезних птахів, що, мабуть, зайняли екологічну нішу великих динозаврів. Однією з них була Diatryma, виявлена у Вайомінгу, зріст 1,8-2,1 м, з масивними ногами, потужним дзьобом і дуже маленькими, недорозвиненими крилами.
У крейдяному періоді жили ящіри або птерозаври, що літають,
Розмах крил вони мали 7,5 м; вони мешкали в Європі, Африці, Північній та Південній Америці і були м'ясоїдними (їли рибою та водними безхребетними).
Великі доісторичні птахи нагадували страусів
 Згідно з палеонтологічними даними, існували великі, частково оперені ящіри. 1834 року французький мандрівник Гудо знайшов на Мадагаскарі половину яєчної шкаралупи такого розміру, що її можна було використовувати як посуд для води. Потім у болотах острова знайшли кілька гігантських кісток, які спочатку прийняли за останки слона чи носорога. Але кістки належали птаху, який мав важити щонайменше півтонни. Мадагаскарські страуси епіорніс (Аepyornithes),досягали висоти 5 м, несли яйця в 32 см завдовжки і 22 см завширшки, що вміщали 8,5 л рідкого вмісту. Найбільшим у кладці епіорнісу вважається яйце довжиною 24 см та об'ємом 11 літрів.
Згідно з палеонтологічними даними, існували великі, частково оперені ящіри. 1834 року французький мандрівник Гудо знайшов на Мадагаскарі половину яєчної шкаралупи такого розміру, що її можна було використовувати як посуд для води. Потім у болотах острова знайшли кілька гігантських кісток, які спочатку прийняли за останки слона чи носорога. Але кістки належали птаху, який мав важити щонайменше півтонни. Мадагаскарські страуси епіорніс (Аepyornithes),досягали висоти 5 м, несли яйця в 32 см завдовжки і 22 см завширшки, що вміщали 8,5 л рідкого вмісту. Найбільшим у кладці епіорнісу вважається яйце довжиною 24 см та об'ємом 11 літрів.
Птах Рух
Венеціанському мандрівнику Марко Поло не довелося самому побувати на Мадагаскарі, але й він чув дивовижні історії: «Розповідають, що там є птах гриф, з'являється у певну пору року, і в усьому гриф не такий, як у нас думають і як його зображають. У нас кажуть, що гриф наполовину птах, наполовину лев, і це неправда. Ті, хто його бачив, запевняють, що він схожий на орла, але дуже великий... Звати його на острові Руком».
Епіорніси були окільцьовані 5 тис. років тому.
 Французькі зоологи виявили на Мадагаскарі останки епіорнісу із бронзовим кільцем, прикріпленим до ноги птиці. Експерти дійшли висновку, що знаки на кільці не що інше, як відбиток печатки епохи найдавнішої цивілізації Індії – Мохенджо-Даро. виготовленої близько п'яти тисяч років тому. Радіовуглецевий аналіз кісток птиці допоміг встановити її вік: він дорівнює п'яти тисячоліть! У третьому тисячолітті до нашої ери жителі Індостану здійснювали сміливі морські експедиції. На той час вони мали багатовіковий досвід водіння кораблів, побували індійці і Мадагаскарі. Тоді тут удосталь водилися епіорніс. В оповіданнях мореплавців, що повернулися додому, їм приділялося багато уваги.
Французькі зоологи виявили на Мадагаскарі останки епіорнісу із бронзовим кільцем, прикріпленим до ноги птиці. Експерти дійшли висновку, що знаки на кільці не що інше, як відбиток печатки епохи найдавнішої цивілізації Індії – Мохенджо-Даро. виготовленої близько п'яти тисяч років тому. Радіовуглецевий аналіз кісток птиці допоміг встановити її вік: він дорівнює п'яти тисячоліть! У третьому тисячолітті до нашої ери жителі Індостану здійснювали сміливі морські експедиції. На той час вони мали багатовіковий досвід водіння кораблів, побували індійці і Мадагаскарі. Тоді тут удосталь водилися епіорніс. В оповіданнях мореплавців, що повернулися додому, їм приділялося багато уваги.
Чи існують епіорніс і понині?
Яйця, які знаходили на піщаних дюнах та в болотах у південній частині острова Мадагаскар, виглядали підозріло свіжими. Здавалося, вони знесені нещодавно. Місцеві жителі впевнені, що в найдрішучіших лісах острова досі живуть гігантські птахи, проте побачити їх нелегко. На Мадагаскарі залишилися ще величезні ділянки заповідних джунглів та нехожених боліт, місця для епіорнісу достатньо.
Страус дромоміс з Австралії
Судячи з копалин ніг, знайдених в 1974 р. поблизу Еліс - Спрінгса, нелітаюча Dromomis stirtoni,гігантський, що нагадував страуса птах, який мешкав у центральній частині Австралії в період від приблизно 15 млн. і до 25 000 років тому, досягав у висоту 3 м і важив близько 500 кг.
Cтраус моа з Нової Зеландії
Гігантський птах, схожий на страуса, моа (Dinornis maximus),яка існувала на островах Нової Зеландії імовірно до початку ХIХ століття, мала, ймовірно, ще більший зріст - 3,7 м, а важила близько 230 кг.
Коли вимерли останні гігантські птахи Австралії
Аналіз давньої яєчної шкаралупи дає підстави припустити, що величезні птахи Австралії, що не літають, вимерли 45-55 тисяч років тому через те, що люди випалили їхнє середовище проживання.
Що їли перші птахи
Міжнародна група вчених провела аналіз сотень фрагментів яєчної шкаралупи зниклих птахів, що не літали, які називалися геніорніс і жили 130-50 тисяч років тому. Вуглецеві ізотопи яєчної шкаралупи показують, що їли птахи, коли відкладали яйця. Виявилося, що раціон геніорнісу був суворим і завжди включав траву.
Найбільший древній птах, здатний до польоту
 Наприкінці третинного періоду (1 млн. років тому) і протягом раннього плейстоцену, або епохи заледеніння, чисельність та різноманітність птахів досягли максимуму. З'явилося багато нинішніх видів, а також інші види, які згодом вимерли. Teratornis incredibilisз Невади (США), величезний нагадував кондора птах з розмахом крил 4,8-5,1 м; ймовірно, була найбільшою з відомих пернатих, здатних до польоту.
Наприкінці третинного періоду (1 млн. років тому) і протягом раннього плейстоцену, або епохи заледеніння, чисельність та різноманітність птахів досягли максимуму. З'явилося багато нинішніх видів, а також інші види, які згодом вимерли. Teratornis incredibilisз Невади (США), величезний нагадував кондора птах з розмахом крил 4,8-5,1 м; ймовірно, була найбільшою з відомих пернатих, здатних до польоту.
Відмінність птахів від ссавців
Ознаки, специфічні для класу птахів, пов'язані насамперед із здатністю цих тварин до польоту, хоча деякі їх види, наприклад страуси та пінгвіни, у ході своєї пізнішої еволюції її втратили. Ще сильніше вони виділяються завдяки своїм пір'ям, які не виявлені в жодної іншої тварини. Від більшості ссавців вони відрізняються тим, що відкладають яйця.
Зниклі та зникаючі птахи
 Перший зафіксований документально випадок такого роду - знищення дронта. Маврикійські дронти Raphus cuculatusвеликі голуби, що не летять, на вигляд нагадують індиків, три види яких мешкали на трьох островах Маскаренського архіпелагу в Індійському океані (Маврикії, Реюньоні і Родрігесі). Вони були стрімко знищені людством практично відразу після їх відкриття: архіпелаг було відкрито 1507 року, останнього дронта на Маврикії бачили 1681 року. За 174 року після відкриття Маврикія європейцями в 1507 р. вся популяція цих птахів була винищена моряками і тваринами, яких вони завезли на своїх кораблях. На острові Реюньоні останнього птаха вбили в 1750 році, на острові Родрігес останній птах теж не дожив до кінця 18 століття.
Перший зафіксований документально випадок такого роду - знищення дронта. Маврикійські дронти Raphus cuculatusвеликі голуби, що не летять, на вигляд нагадують індиків, три види яких мешкали на трьох островах Маскаренського архіпелагу в Індійському океані (Маврикії, Реюньоні і Родрігесі). Вони були стрімко знищені людством практично відразу після їх відкриття: архіпелаг було відкрито 1507 року, останнього дронта на Маврикії бачили 1681 року. За 174 року після відкриття Маврикія європейцями в 1507 р. вся популяція цих птахів була винищена моряками і тваринами, яких вони завезли на своїх кораблях. На острові Реюньоні останнього птаха вбили в 1750 році, на острові Родрігес останній птах теж не дожив до кінця 18 століття.
Найвідоміші з вимерлих птахів

Мандрівний голуб
У 1914 р. у зоопарку міста Цинциннаті (Північна Кароліна, США) загинула Марта – остання представниця раніше численного роду мандрівних голубів (Ectopistes migratorius).Мандрівні голуби були безжально винищені на м'ясо.
Першим вимерлим від рук людини видом Північної Америки
…стала безкрила гагарка (Alca impennis),вимерла в 1844 році. Вона також не літала і гніздилася колоніями на атлантичних островах поблизу континенту. Матроси та рибалки легко вбивали цих птахів заради м'яса, жиру та виготовлення наживки для тріски.
Незабаром після зникнення безкрилої гагарки жертвами людини стали два види на сході північноамериканського континенту. Одним із них був каролінський папуга (Conuropsis carolinensis).Фермери у великій кількості вбивали цих зграйних птахів, оскільки тисячі їх регулярно робили набіги на сади.
Зникло 100 видів птахів
З 1600 року у всьому світі зникло, ймовірно, близько 100 видів птахів. Більшість їх було представлено невеликими популяціями на морських островах. Часто не здатні до польоту, як дронт, і люди, що майже не боялися, і привезених їм дрібних хижаків, вони стали легкою їх здобиччю.
Багато видів птахів також знаходяться на межі вимирання В даний час багато видів птахів також знаходяться на межі вимирання або, у кращому випадку, мають його загрозу. У Північній Америці в найгіршому становищі знаходяться каліфорнійський кондор, жовтогий зуйок, американський журавель, ескімоський кроншнеп і білоклювий королівський дятел (можливо вже вимерлий). В інших регіонах велика небезпека загрожує бермудському тайфуннику, філіппінській гарпії, какапо (совиному папузі) з Нової Зеландії – нічному вигляду, що не літає, а також австралійському земляному папузі.
Птахи, яким загрожує вимирання
У природних умовах у наш час живе лише один блакитний ара (Cyanopsittaspixii),проте приблизно 30 таких птахів утримуються в неволі.
Гавайський співочий птах, лускокрилий мохо (Moxobracattus),вважалася повністю вимерлою і вдруге відкрита лише 1960 р., очевидно, представлена лише двома парами особин.
У світі залишилося живих менше 20 (головним чином у неволі) червононогих ібісів (Nipponia nippon), але всі вони, мабуть, занадто старі, щоб розмножуватися.
Внаслідок неконтрольованого полювання новозеландський совиний папуга (Strigops habroptilus)опинився на межі вимирання. Друга причина зникнення полягає в тому, що цей нелітаючий птах не може рятуватися від хижаків, тому в живих залишається всього 10 екземплярів.
Зараз у природних умовах існують лише кілька каліфорнійських кондорів, вирощених у неволі та випущених у 1992 р.
До інших відомих вимерлих видів птахів відносяться

Лабрадорська гага Camptorhynchus labradorius.
Самоанська очеретяниця Gallinula pacifica.
Біла султанка Porphyrio Albus.
Маврикійський синій голуб Alectroenas nitidissima.
Норфолкський земляний голуб Hemiphaga argetraca.
Тонкоклювий нестор Nestor productus.
Кубинський ара Ara tricolor.
Кайманський синій дрізд Turdus ravidus.
У незавидному положенні перераховані вище птахи виявилися головним чином з вини людини, яка поставила їх популяції на межу зникнення безконтрольним полюванням, непродуманим використанням пестицидів або корінним перетворенням природних житла.
На межі зникнення зараз знаходяться 26 видів птахів та 132 види ссавців.
Всім давно відомо, що птахи походять від динозаврів, але мало хто знає про древніх птахів мезозойської ери, що жили пліч-о-пліч з гігантськими рептиліями, або кайнозойської ери - простір геологічного часу після вимирання динозаврів. У наступних 10 пунктах статті наведено опис та фото вимерлих видів доісторичних птахів (у хронологічному порядку їх появи), які справляли значний вплив на еволюційний розвиток пернатих тварин.
1. Aurornis (160 мільйонів років тому)
Можливо, хтось думав, що першим у списку має бути археоптерикс, але невеликі птахи роду aurornisз'явилися раніше за археоптерикс на 10 мільйонів років. Тим не менш, aurornis мав більше спільного з динозаврами, ніж з птахами, а його пір'я було занадто тонким, щоб бути корисним у польоті. Але все ж таки, ми вважатимемо aurornisнайдавнішим птахом, і залишимо суперечки з роздумами палеонтологам.
2. Конфуціусорніси (130 мільйонів років тому)

На відміну від попереднього представника копалин птахів, конфуціусорніси більше схожі на сучасних птахів. Це були перші птахи, що мали справжній дзьоб. Вони не мали зубів (головна риса рептилій), тіло було вкрите товстим шаром пір'я, а довгі, вигнуті пазурі, давали можливість впевнено сидіти на гілках високих дерев. Незважаючи на вищеописане, не можна зробити ствердний висновок, що всі сучасні птахи походять від конфуціусорнісів: ймовірно, що птахи з'являлися і вимирали незалежно один від одного кілька разів протягом мезозойської ери.
3. Gansus (110 мільйонів років тому)

Як ви вже зрозуміли з перших пунктів, дуже складно (або навіть неможливо) повністю розібратися в еволюції птахів, які жили десятки мільйонів років тому. Gansus- ще один вид доісторичних птахів, який подається в палеонтологічних колах, як найдавніший представник підкласу справжніх птахів (тобто прямий предок усіх сучасних птахів). Ця теорія викликає безліч суперечок, але все ж таки, давно вимерлий gansusнайкращий претендент як прабатька сучасних качок та гагар.
4. Гесперорніси (75 мільйонів років тому)

Стародавні птахи мали достатньо часу для розвитку та дееволюції у другій половині мезозойської ери. Цікавим є той факт, що птахи роду гесперорніс були вдруге налітаючими (тобто вони еволюціонували з ранніх літаючих птахів, але поступово втратили здатність літати, як пінгвін або індичка). Можливо, це сталося через конкуренцію з великими птерозаврами пізнього крейдяного періоду Північної Америки, зокрема всюдисущих птеранодонів, тому гесперорнісам довелося задовольнятися наземною екологічною нішою.
5. Гасторніси (55 мільйонів років тому)

Після загибелі динозаврів, близько 65 мільйонів років тому, птахи змогли розвиватися в екологічних нішах, що звільнилися. Роль жахливого двоногого хижака, перейшла до двох метрових птахів з роду гасторніс (також відомих, як діатріми). Передбачається, що гасторніс полювали в зграях, переслідуючи своїх жертв наче зменшені копії тиранозавра рекс.
6. Eocypselus (50 мільйонів років тому)

Ви колись запитували, як виглядав доісторичний предок колібрі? Палеонтологи особливо не поширюються з цього приводу, але все ж таки давно відомо, що колібрі походять від eocypselus- Вигляд дрібних птахів, що мешкають у лісовій місцевості раннього еоцену Північної Америки, близько 50 мільйонів років тому. Крила eocypselusбули більшими, ніж у сучасного колібрі, тому його політ не можна було назвати граціозним.
7. Icadyptes salasi - предок пінгвінів (40 мільйонів років тому)

Можна припустити, що стародавні пінгвіни, що жили близько 40 мільйонів років тому мали такий самий спосіб життя, як і сучасні: жили на крижинах, пірнали за рибою і при кожній нагоді чистили пір'я. Значною мірою це припущення вірне, за винятком життя на льоду. Наприкінці еоцену, icadyptesнасправді мешкали у тропічному кліматі поблизу екватора Південної Америки. Ці пінгвіни були більшими за сучасні види і досягали 1,5 м у висоту і маси близько 35 кг.
8. Фороракоси (12 мільйонів років тому)

Пам'ятаєте гасторніс (див. пункт 6), 2 м заввишки та масою понад 100 кг, які жили протягом десяти мільйонів років після динозаврів? Так ось, фороракоси через 40 мільйонів років стали гідною заміною гасторнісів. Значною мірою фороракоси вели той самий спосіб життя, що й гасторніс. Хоча, у них була додаткова зброя в арсеналі: довга, потужна наче сокира дзьоба, яку вони використовували, щоб завдати глибоких смертельних ран своїм жертвам.
9. Аргентавіс (6 мільйонів років тому)

Хоч би як вражаюче виглядали птахи протягом кайнозойської ери, вони ніколи не відповідали розмірам і величі найбільших птерозаврів. Наприкінці епохи міоцену, аргентавіс був найбільшим літаючим птахом, маючи розмах крил до 7 м і масу 70-72 кг. Вражає? Але, 60 мільйонів років до цього, птерозавр кетцалькоатль мав розмах крил близько 12 м (як у приватного літака). Як не дивно, менш габаритний аргентавіс літав подібно до птерозаврів, також ширяючи на повітряних потоках, а не активно махаючи крилами.
10. Епіорнісові (2 мільйони років тому)

Епоха плейстоцену, від 2 мільйонів до 10 тисяч років тому, стала періодом повернення мегафауни. Крім шаблезубих тигрів і мамонтів, плейстоцен подарував гігантські птахів, таких як мадагаскарський епіорніс із сімейства епіорнісових. У висоту ці птахи досягали 3-5 м і мали масу тіла до 500 кг, які яйця приблизно 100 разів перевищували обсяг стандартного курячого яйця.
Наука
Легендарна крилата істота, досі відома як найперший птах на землі – археоптерикс, швидше за все, буде позбавлений такого звання. Натомість, на основі нещодавно виявлених копалин у Китаї було висунуто припущення, що археоптерикс був зовсім не птахом, а лише одним із багатьох видів "пташиних" динозаврів.Дане відкриття може змусити вчених переглянути багато з того, що вони знали про походження та еволюцію птахів.
Археоптерикс жив приблизно 150 мільйонів років тому на території нинішньої Баварії (Німеччина), коли Європа була острівним архіпелагом у теплому мілководді тропічних морів. Вперше виявлені 150 років тому, викопні останки із сумішшю пташиних рис та характеристик рептилій, здавалося, закріплюють і пояснюють еволюційний зв'язок між динозаврами та птахами, тим самим підтверджуючи теорію еволюції Дарвіна, опубліковану за два роки до цього. Отримавши звання найпершої та найпримітивнішої птиці на землі, археоптерикс став центральною фігурою в розумінні вченими еволюції птахів та польоту.
Тим не менш, протягом десятиліть все ж таки існували сумніви в тому, чи був він насправді птахом. "За останні 15-20 років, здавалося б, такі однозначно пташині характеристики археоптериксу, як пір'я, грудна кістка та трипала кінцівка, відкрилися вченим з іншого погляду, тобто вони, як з'ясувалося, не властиві лише царству птахів", - каже анатом. та палеонтолог університету штату Огайо Лоуренс Вітмер (Lawrence Witmer). "Ми почали знаходити велику кількість динозаврів з пір'ям, причому багато з них мали добре розвинену грудну кістку, як у археоптерикса" , – продовжує він.
В даний час вчені виявили нові копалини в Китаї, поєднання характеристик яких несподівано говорить про те, що археоптерис був просто однією з ланок того ланцюжка, який зрештою сприяв появі птахів. Виявлене розміром з курку викопне Xiaotingia zhengi віком близько 155 мільйонів років. Останки м'ясоїдної тварини знайшли в провінції Ляонін (Liaoning) у Китаї, де раніше знаходили й інші ексклюзивні зразки багатьох пернатих динозаврів.
Для того, щоб зрозуміти до якого виду еволюційно належить знайдені викопні останки, палеонтолог при китайській академії наук у Пекіні Ксін Ксу (Xing Xu) та його колеги співвіднесли всі присутні у знахідки пташині риси та властивості динозавра з іншими видами.Серед цих видів були і авіалани (предки сучасних птахів), і дейноніхозаври, близькі родичі авіаланів, разом відомі як паравіани. До групи авіаланів входять динозавро-подібні птахи, тоді як дейноніхозаври – це птахи динозаври. Однак, дуже розмиті відмінності між групами призвели до того, що багато вчених вирішили відмовитися від поділу їх на такі типи.
Коли вчені вирішили проаналізувати особливості археоптерикса і порівняти його з нещодавно знайденими останками, вони в результаті дійшли висновку, що об'єкти, що порівнюються, належать до однієї групи.
Таким чином, зараз отримані дані говорять про те, що найраніші з відомих авіаланів – це розміром з голуба перната істота, яка називається епідексиптерикс ху (Epidexipteryx hui), останки якої були нещодавно виявлені в Китаї. "Отримані дані мають великий вплив на наші уявлення про те, що ми вважали ранньою еволюцією птахів. За 150 минулих років вчені розглядали еволюцію крізь призму археоптерикса, скільки ж інформації їм тепер доведеться переглянути? - запитує Вітмер, який брав участь у дослідженні.
Проте результати поки що залишаються попередніми. "Нові знахідки змінюють наше бачення та розуміння багатьох речей, проте це не означає, що якщо сьогодні виявлені останки Xiaotingia прибрали археоптерикса з розряду птахів, завтра ми не знайдемо щось ще, що зможе повернути його назад", - підсумував Вітмер.