Жлези и техните хормони в тялото
В човешкото тяло освен основните функционални органи има и така наречените жлези, тоест органи, предназначени да синтезират и секретират активни вещества, наречени тайни. Жлезите често са микроскопични и само няколко са относително големи. Интересното е, че жлезите, които имат отделителни канали и секретират тайна за лигавицата или повърхността на тялото (пот, слюнка, мляко и други), се наричат \u200b\u200bекзокринни, а нямат канали и отделят високоактивни вещества (хормони) директно в кръвоносната система (щитовидна жлеза, надбъбречни жлези , хипофизната жлеза и други) се наричат \u200b\u200bендокринни. Жлезите и техните хормони трябва да бъдат разгледани по-подробно.
Разположен пред ларинкса на врата на човек, той произвежда хормони, богати на йод - тироксин, трийодтиронин и калцитонин. Тези хормони са предназначени да регулират въглехидратния, мастния и протеиновия метаболизъм в организма, а освен това влияят върху растежа и умственото развитие на човека. С хиперфункция на тази жлеза човек развива затлъстяване, телесната температура се повишава и метаболизмът намалява.
Те са малки образувания над бъбреците, състоящи се от два слоя от външния (кортикален), както и от вътрешния (мозъчен). Външният слой произвежда полови хормони, които са отговорни за развитието на половите характеристики и регулират метаболизма (протеини и мазнини, въглехидрати, калий, натрий). При дисфункция на надбъбречната кора човек се разболява, така наречената бронзова болест, тоест тялото му придобива бронзов цвят, а самият той изпитва постоянна умора, загуба на апетит и гадене. Функцията на надбъбречната медула включва производството на хормони като норепинефрин и адреналин. И така, адреналинът увеличава сърдечната честота и ускорява кръвообращението, мобилизирайки всички сили на тялото вътре стресова ситуация... Не липсва този хормон, но излишъкът му заплашва със свиване на кръвоносните съдове и учестен пулс. Напротив, норепинефринът е създаден да забавя сърдечната честота.
Панкреасът, разположен в коремната кухина, е жлеза със смесен секрет, тъй като има отделителни канали и отделя секрети в стомаха, като участва в храносмилането. Един от хормоните, които произвежда, гликогенът синтезира глюкоза, като по този начин повишава нивата на кръвната захар. Другият хормон, инсулинът, напротив, е единственият, който понижава нивата на кръвната захар. Именно поради дисфункцията на панкреаса диабет, при които има повишаване на кръвната захар.
Половите жлези, тоест женските яйчници и мъжките тестиси, също са жлези със смесена секреция. Поради външната си секреция се появяват зародишни клетки, тоест сперматозоиди и яйца, а тяхната ендокринна функция е да произвеждат мъжки, както и женски полови хормони. Тези хормони влияят на сексуалното поведение и фигурата на човек от определен пол. Мъжките тестиси произвеждат андрогени, които са отговорни за наличието на мъжки сексуални характеристики, мускули, мустаци, брада, а женските яйчници произвеждат естрогени, отговорни за половите характеристики на нежния пол, тоест развитието на млечните жлези и формата на тялото.
Хипофизната жлеза, разположена в мозъка, се състои от 3 дяла (преден, междинен и заден), два от които произвеждат хормони. В предния лоб се произвежда хормон, който влияе върху растежа на костите и ускорява метаболитните процеси. Недостигът му може да застраши развитието на нанизъм, а хиперфункцията може да доведе до гигантизъм. Междинната хипофизна жлеза произвежда хормон, отговорен за пигментацията на кожата.
Всичко по-горе само потвърждава, че жлезите и техните хормони са изключително важни за човешкото тяло и без тях човек не би могъл да съществува. Бягай сам!
Популярни в сайта
Информацията на нашия уебсайт има информативен и образователен характер. Тази информация обаче по никакъв начин не е предназначена за ръководство за самолечение. Не забравяйте да се консултирате с Вашия лекар.
За да се разбере как функционират ендокринните органи, и по-специално щитовидната жлеза, е необходимо накратко да се разгледа механизмът на действие на хормоните.
Фигура: 1. Разположението на ендокринните органи
Ендокринната функция на тялото се осигурява от системи, които включват:
1) жлези с вътрешна секреция, които отделят хормони;
2) хормони и различни начини за транспортирането им;
3) подходящи органи или целеви тъкани, които реагират на действието на хормоните.
Ендокринната система поддържа постоянството на вътрешната среда на тялото, което е необходимо за нормалното протичане на физиологичните процеси.
Ендокринните жлези са специализирани органи с жлезиста структура. Има жлези само с вътрешна секреция (хипофиза, надбъбречни жлези, щитовидна жлеза, паращитовидни жлези) и смесени - с вътрешна и външна секреция. Пример за това е панкреасът. Външната му секреция се състои в производството на храносмилателни ензими, които чрез специален канал навлизат в дванадесетопръстника, а вътрешната секреция се състои в производството на хормона инсулин в специализираните бета-клетки на панкреатичните островчета (Лангерханс), който отива директно в кръвта и регулира кръвната захар ... Половите жлези също извършват вътрешни и външни секрети.
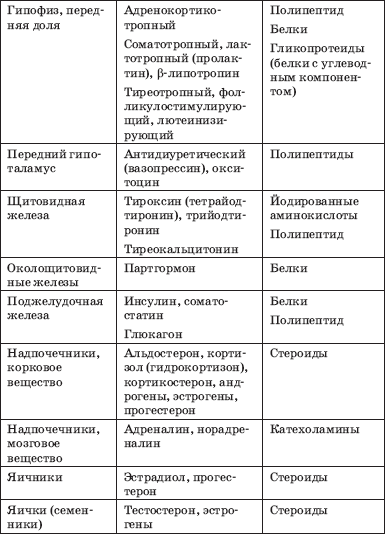
Името и местоположението на жлезите с вътрешна секреция, хормоните, които те произвеждат, химичното естество на последните са представени в табл. 1.
Таблица 1. Хормони на жлезите с вътрешна секреция (Potemkin V.V., 1986)

Краят на таблицата. 1
Терминът "хормон", преведен от гръцки, означаващ "възбуждам", "предизвиквам", е въведен на практика от Bayliss и Starling. През януари 1902 г. те провеждат своя известен, класически експеримент, който убедително доказва участието на хуморалния фактор в регулирането на секреторната дейност на панкреаса. Бейлис и Старлинг считат за хормон всяко вещество, което обикновено се произвежда от клетките на която и да е част от тялото и се пренася от кръвта в отдалечени части, върху които той действа в полза на тялото като цяло.
В момента хормоните се определят като високоактивни вещества, образувани в жлезите с вътрешна секреция, навлизащи в кръвния поток и упражняващи регулаторен ефект върху функциите на органите и телесните системи, отдалечени от мястото на секрецията им. Те се наричат \u200b\u200bоще химически пратеници, които се секретират директно в кръвта от специализирани клетки, които са в състояние да синтезират и освобождават хормони в отговор на специфични сигнали.
Според своята химическа структура хормоните се разделят на:
1) хормони - производни на аминокиселини;
2) протеинови и полипептидни хормони;
3) стероидни хормони.
Според своя физиологичен ефект хормоните се разделят на тригери и изпълнители. Изходните хормони (активатори на дейността на други ендокринни жлези) включват неврохормоните на хипоталамуса и тропните хормони на хипофизната жлеза. Изпълняващите хормони имат пряк ефект върху основните функции на тялото.
Хормоните се различават от другите биологично активни вещества по следните свойства:
1) много висока биологична активност;
2) отдалечения характер на действието;
3) строга специфичност.
Високата биологична активност на хормоните се характеризира с факта, че, намирайки се в кръвта в незначителни количества, те имат подчертан ефект.
Дистанционният характер на действието на хормоните се крие във факта, че точките на приложение на тяхното действие обикновено се намират далеч от мястото на образуване на хормона в ендокринната жлеза.
Хормоните се отличават със строга специфичност на действието. Това означава, че реакциите на органи, тъкани и клетки към хормони са строго селективни. Всеки хормон действа само в определени органи и тъкани, така наречените целеви органи (целеви тъкани). Хормонът разпознава и взаимодейства със своя целеви орган, защото в тези органи има специални съединения - рецептори. Рецепторите са информационни протеинови молекули, които разпознават и трансформират хормоналния сигнал в хормонално действие. Към днешна дата са идентифицирани повече от 60 рецептора. За стероиди (хормони на кората на надбъбречната жлеза) и хормони на щитовидната жлеза (хормони щитовидната жлеза), които лесно проникват през мембраната, рецепторните протеини се намират вътре в клетката. Рецепторите за протеинови хормони и катехоламини, които не могат да преминат през клетъчната мембрана, са разположени на клетъчната повърхност.
Хипоталамусът и хипофизната жлеза са единна система за контрол на периферните ендокринни жлези.
Хипоталамусът е част от мозъка, която има свойствата на нервната и ендокринната системи. Хипоталамусът получава обширен поток от информация от сетивата и вътрешните органи. Съставът на невросекреторните ядра на хипоталамуса включва така наречените едро-клетъчни и дребноклетъчни ядра. Първите секретират хормоните окситоцин и вазопресин, които се транспортират по нервните стволове до задния лоб на хипофизната жлеза, натрупват се там и при необходимост се използват за регулиране на дейността на бъбреците и матката.
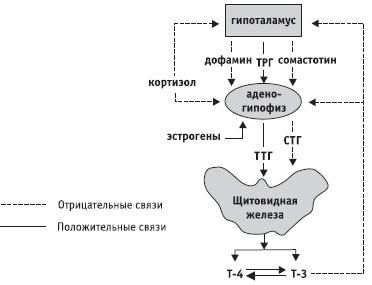
Фигура: 2. Схема на регулиране на хипоталамо-хипофизно-щитовидната система
Други функции се изпълняват от малките клетъчни ядра на хипоталамуса. Те са способни да произвеждат така наречените освобождаващи хормони или, по-правилно, освобождаващи фактори (разрешаващи фактори). Освобождаващите фактори чрез венозната система достигат до хипофизната жлеза и регулират отделянето на хормони на последната.
Регулирането на дейността на хипофизната жлеза от хормоните на дребноклетъчните ядра на хипоталамуса се извършва съгласно антагонистичния принцип. Едната група фактори стимулира отделянето на хипофизни хормони (освобождаващи фактори или либерини), а другата инхибира (статини). Известни са следните фактори: кортиколиберин, който стимулира секрецията на адренокортикотропен хормон на хипофизната жлеза; тиролиберин, който засилва отделянето на тиреоид-стимулиращ хормон от хипофизната жлеза; соматолиберин и соматостатин (първият стимулира отделянето на хипофизния соматотропен хормон - хормон на растежа, а вторият инхибира); меланолиберин и меланоцитостатин и др.
Хипофизната жлеза е централната ендокринна жлеза, в която се произвеждат така наречените тропични хормони, които регулират функцията на периферните жлези. Това е сложен ендокринен орган, разположен в основата на мозъка - в така нареченото турско седло. Състои се от аденохипофизата, по-голямата част от която е предният лоб на жлезата, и неврохипофизата, представена от задния й лоб.
В предния лоб (аденохипофиза) се произвеждат тропични хормони:
Хормон на растежа, който регулира растежа на тялото, синтеза на протеини, разграждането на глюкозата и мазнините;
Кортикотропин, който стимулира синтеза на глюкокортикоиди в надбъбречната кора;
Тиротропин - стимулатор на синтеза на тиреоидни хормони на щитовидната жлеза;
Гонадотропин, фоликулотропин, регулиращ синтеза на мъжки и женски полови хормони;
Пролактинът е хормон, който регулира лактацията.
В задния лоб на хипофизната жлеза (неврохипофиза) се натрупват вазопресин и окситоцин. Вазопресин, или антидиуретичен хормон, регулира водния обмен и съдовия тонус. Окситоцинът повишава тонуса на гладката мускулатура на матката, регулира раждането и отделянето на мляко от млечните жлези.
Периферните жлези с вътрешна секреция са разделени на две групи.
Първата се състои от жлези, чиято функция се регулира от тропните хормони на аденохипофизата. Те се наричат \u200b\u200bаденохипофизозависими жлези или целеви жлези. Те включват щитовидната жлеза, кората на надбъбречната жлеза и ендокринните части на половите жлези. Връзката между аденохипофизата и целевите жлези се основава на принципа на „обратна връзка“. Например аденохипофизата секретира тиреостимулиращ хормон в кръвния поток, който стимулира секрецията на тиреоидния хормон тироксин. Тироксинът, влязъл в кръвта, инхибира освобождаването на тиреоид-стимулиращ хормон от хипофизната жлеза.
Втората група периферни ендокринни жлези се състои от жлези, чиято функция не зависи от дейността на хипофизната жлеза. Тези жлези се наричат \u200b\u200bзависими от аденохипофиза. Те функционират офлайн. Те включват паращитовидните жлези, ендокринната част на панкреаса, надбъбречната медула и ендокринните клетки на тимусната жлеза.
Тимусът (тимус или тимус, жлеза) произвежда хормоните тимозини и тимопоетини, които стимулират имунните процеси.
Щитовидна жлеза произвежда йодсъдържащи хормони: тироксин и трийодтиронин, както и тирокалцитонин. Тироксинът и трийодтиронинът регулират основния метаболизъм, тоест нивото на консумация на енергия, необходимо за поддържане на жизнените функции на организма в състояние на пълноценен покой. Тирокалцитонинът регулира обмена на калций и фосфор.
В паращитовидните жлези се произвежда паратиреоиден хормон, който също регулира метаболизма на калция и фосфора. Но ако тиреокалцитонинът на щитовидната жлеза понижава съдържанието на калций в кръвта, тогава паращитовидният хормон го увеличава. Антагонистичната връзка между тирокалцитонин и паратиреоиден хормон осигурява нивото на калций в кръвта на нивото, необходимо за организма.
Ролята на надбъбречните хормони е изключително важна. Това са сдвоени органи, разположени над горните полюси на бъбреците. В надбъбречните жлези се разграничават кората и медулата.
Корковото вещество секретира група стероидни хормони, наречени заедно кортикостероиди. Три зони на кората са специализирани за секрецията на различни хормони. Клетките на гломерулната зона произвеждат минералокортикоиди: дезоксикортикостерон и алдостерон, които регулират минералния метаболизъм. Зоната на снопа произвежда глюкокортикоиди: кортизол и кортикостерон, които регулират метаболизма на протеини, мазнини и въглехидрати. В ретикуларната област се синтезират някои предшественици на мъжки полови хормони (андрогени).
Медулата на надбъбречната жлеза освобождава катехоламини - адреналин и норепинефрин - в кръвта. Норадреналинът действа не само като хормон, но и като медиатор на нервните процеси симпатично разделение нервна система... Катехоламините имат подчертан вазоконстрикторно действие, като по този начин повишават кръвното налягане. Те участват в регулирането на въглехидратния и мастния метаболизъм, играят основна роля в адаптацията на тялото по време на стрес. Адреналинът се освобождава в отговор на голямо разнообразие от стимули: страх, вълнение, болка, радост. Образно се нарича хормон на спешността, хормонът на емоциите, първият медиатор на стреса.
В ендокринната част на панкреаса (островчета на Лангерханс) се произвеждат инсулин, глюкагон и соматостатин. Инсулинът е най-важният регулатор на въглехидратния, както и метаболизма на мазнините и протеините. Глюкагонът е физиологичен инсулинов антагонист, както и стимулатор на секрецията на инсулин в присъствието на глюкоза. Соматостатинът инхибира секрецията на инсулин, глюкагон и хормон на растежа. Нарушената секреция на инсулин и глюкагон води до развитието на такова сериозно и широко разпространено заболяване като захарен диабет.
Половите жлези произвеждат не само хормони, но и полови клетки (сперматозоиди и яйцеклетки). Тестисите (тестисите) произвеждат мъжки полови хормони - андрогени, основният от които е тестостеронът. Андрогените допринасят за развитието на първични и вторични мъжки полови характеристики. Яйчниците синтезират женски полови хормони - естрогени, които са отговорни за образуването на женски първични и вторични полови белези, както и прогестерон, хормон, необходим за нормалното протичане на бременността. Производството на хормони и зародишни клетки се извършва под контрола на гонадотропните хормони на аденохипофизата.
Бъбреците, изпълняващи отделителна функция, също са вид ендокринна жлеза. Клетките на така наречения юкстагломеруларен апарат на бъбреците секретират в кръвта хормона ренин, който участва в образуването на ангиотензин II - активен регулатор на съдовия тонус. Бъбреците също произвеждат еритропоетин, хормон, който стимулира образуването на червени кръвни клетки в костния мозък.
Установено е, че сърцето също е ендокринна жлеза. В атриума се синтезира натриуретичен хормон, който влияе върху екскрецията на натрий през бъбреците.
Временно функциониращият ендокринен орган е плацентата („място на бебето“). Той произвежда хормони, които допринасят за нормалния ход на бременността.
В централната нервна система се образуват специални вещества - невроендокринни пептиди (неврохормони) - ендорфини, енкефалини. Те се наричат \u200b\u200b"ендогенни опиати" или морфиноподобни пептиди. Тези хормони имат аналгетични (облекчаващи болката) ефекти и възпроизвеждат поведенческите ефекти на морфина.
Единството и взаимовръзката на нервните и ендокринните механизми е много ясно проследено в примера за функционирането на хипоталамо-хипофизната система. Понастоящем е по-правилно да се говори не за ендокринната система, а за невроендокринната система на тялото.
След като очертахме общите идеи за жлезите с вътрешна секреция, нека преминем към основната цел на нашата история - щитовидната жлеза.
Въпрос номер 2 Ендокринни жлезии техните хормони.
Ендокринни жлези(от гръцки endon - вътре, crio - разпределям) или жлези с вътрешна секреция, са специализирани органи или групи клетки, чиято основна функция е да развиват и освобождават специфични биологично активни вещества във вътрешната среда на тялото. Ендокринните жлези нямат отделителни канали. Клетките им са плетени от изобилна мрежа от кръвоносни и лимфни съдове, а отпадъчните продукти се отделят директно в кръвта, лимфата и тъканната течност. Тази характеристика фундаментално отличава жлезите с вътрешна секреция от жлезите с външна секреция, които отделят секрета си през отделителните канали.
Продуктите, произведени от жлезите с вътрешна секреция, се наричат хормони(Гръцки hormao - възбуждам, активирам). Терминът "вътрешна секреция" е предложен през 1885 г. от френския физиолог C. Bernard, а терминът "хормон" - от английските физиолози W. Beilis и E. Starling през 1902 г.
Следните характеристики са характерни за хормоните: Техният синтез и изолиране се извършва от специализирани клетки. Хормоните се образуват в жлезистите ендокринни клетки, след което те навлизат във вътрешната среда, главно в кръвта и лимфата. Междинните продукти на синтеза или метаболизма на хормоните често имат биологична активност, но като правило те не са серкетирани .
Висока биологична активност.Хормоните имат физиологичен ефект при много ниски концентрации. И така, концентрацията на женския полов хормон (естрадиол) в кръвта варира от 0,2 до 0,6 μg (10 -6 g) в 100 ml плазма. Съдържанието на растежен хормон в кръвта се измерва в още по-малки количества - нанограми (KG 9 g). Хипофизната жлеза реагира на пикограми (10 -12 g) хипоталамусни хормони, ангиотензин-Р-продукт на ендокринни бъбречни клетки - предизвиква чувство на жажда при фемтограми (10 -15 g). Освен хормоните, никакви други химически отпадъчни продукти не са ефективни в такива малки дози.
Специфичност.Всеки хормон се характеризира с определена, присъща само на него химическа структура, място на синтез и функция. В тази връзка дефицитът на който и да е хормон не може да бъде попълнен с други хормони или биологично активни вещества.
Действие от разстояние.Хормоните, като правило, се носят с кръв далеч от мястото на образуване, засягайки отдалечени органи и тъкани. По този начин те се различават от медиаторите и цитокините, действащи върху една клетка или група клетки на мястото на тяхното образуване.
Химичната структура на хормоните и технитетрансформация в тялото
Според своята химическа структура хормоните могат да бъдат разделени на 4 основни групи: протеини и пептиди, производни на аминокиселини, стероиди, простагландини.
Примери за протеинови хормони: инсулин, соматотропин (хормон на растежа), тропни хормони на предната хипофизна жлеза. Някои от тях (фолитропин, тиротропин, лутропин) са сложни протеини, други (инсулин, калцитонин и др.) Са прости протеини. Пептидната структура има глюкагон, вазопресин, окситоцин, хипоталамусни хормони. Производните на аминокиселини включват хормони на щитовидната жлеза - трийодтиронин, тироксин, както и адреналин и норепинефрин. Стероидните хормони се основават на ядрото на цикличния въглеводород циклопентанперхидрофенантрен. Тази група включва хормони на надбъбречната кора и половите жлези.
Основните етапи на образуването и трансформацията на хормона могат да бъдат представени както следва:
биосинтеза на хормони;
секреция, т.е. изолация от ендокринна клетка;
пренос на кръв към периферните тъкани;
разпознаване на хормоналния сигнал от целевите клетки;
трансдукция (транслация) на хормонален сигнал в биологичен отговор;
потискане на хормоналния сигнал.
Органи и тъкани с клетъчна ендокринна функция
|
|
|
|
Органи и тъкани
|
|
|
|
|
|
Плацента
|
Синцитиотрофобласти Цитотрофобласти
|
Хорион гонадотропин, пролактин Естриол, прогестерон
|
|
|
|
|
Тимоцити
|
Тимозин, тимопоетин
|
|
|
|
|
Интерстициум Тръбички
|
Еритропоетин Калцитриол
|
|
|
|
|
Предсърдни миоцити
|
Атриопептид, соматостатин, ангиотензин-Р
|
|
|
|
|
|
|
|
Органи, тъкани и клетки с ендокринна функция
|
|
|
|
Структури
|
Тъкани, клетки
|
Хормони
|
|
|
|
Хипоталамус
|
Нервни клетки на структури с малка точка
|
Хипоталамусни неврохормони:
|
|
|
|
|
Заден хипоталамус Нервни клетки на големите клетъчни ядра на предния хипоталамус
|
а) либерини: кортиколиберин, тиролиберин, лулиберин, фолиберин, соматолиберин, пролактолиберин, меланолиберин; б) статини: пролактостатин, меланостатин, соматостатин Вазопресин, окситоцин
|
|
|
|
Аденохипофиза Неврохипофиза
|
Кортикотрофи Гонадотрофи Тиротрофи Соматотрофи Лактотрофи Питуицити
|
Кортикотропин, меланотропин Фолитропин, лутропин Тиротропин Растежен хормон Пролактин Вазопресин, окситоцин
|
|
|
|
Надбъбречни жлези Cortex Мозъчна материя
|
Зона на гломерулите Зона на снопчета Мрежова зона Хромафинови клетки
|
Минералокортикоиди Глюкокортикоиди Полови стероиди: андрогени, естрогени Андреналин, норепинефрин
|
|
|
|
Щитовидна жлеза
|
Фоликуларни тироцити
|
Трийодтиронин, тетрайодтиронин Калцитонин, катакалцин, Ко-калцигенин
|
|
|
|
Паращитовидните жлези
|
K-клетъчни основни клетки
|
Паратирин калцитонин
|
|
|
|
|
Пинеоцити
|
Мелатонин
|
|
|
|
Панкреас
|
Островчета Лангерханс: осини клетки R-клетки А-клетки
|
Глюкагон Соматостатин
|
|
|
|
Полови жлези (тестиси, яйчници)
|
Leydig клетки Sertoli клетки Granulosa клетки Жълто тяло
|
Тестостерон естрогени, инхибин естрадиол, естрон, прогестерон Прогестерон
|
|
|
|
|
|
|
|
Биосинтеза на хормонипрограмирани в генетичния апарат на специализирани ендокринни клетки. Следователно, това зависи от структурата и експресията на гени, кодиращи синтеза на тези хормони, както и от ензимите, които регулират хормоналния синтез и пост-транслационните процеси. Липсата или дефектът на съответните гени води до ендокринопатия. Пример за това е нанизъм с генетичен дефект на хормона на растежа.
2. Секреция на хормони.най-важната характеристика на всеки хормон е неговата секреция. Опакован във везикули или гранули, хормонът се транспортира към цитоплазматичната мембрана. Освобождаването на протеин-пептидни хормони и катехоламини от клетката се предшества от взаимодействието на цитоплазматичната мембрана и мембраната на секреторната гранула. След това се случва техният лизис и хормонът се освобождава от клетката. Този процес се активира от много фактори, медиатори, висока концентрация на калий, електрически стимули и др. Хормоналната секреция е акт, придружен от разход на енергия, поради което винаги е свързан с промени в системата АТФ - цАМФ. За секрецията е необходимо участието на калциеви йони, които активират протеините на микротубуларно-микрофиламентната система, улеснявайки взаимодействието на хормонални гранули с тях и влияят върху образуването на цАМФ. Следователно намаляването на съдържанието на калциеви йони в извънклетъчната среда и навлизането му в ендокринната клетка неизбежно води до намаляване на секреторната активност на тази клетка.
3.
Транспорт на хормони.Секретираният хормон навлиза във вътрешната среда на тялото, главно в кръвта, и се транспортира по-нататък. Повечето хормони образуват сложни съединения с плазмени протеини в кръвта. Някои от тези протеини са специфични транспортни протеини (например транскортин, който свързва хормоните на надбъбречната кора), а някои са неспецифични (например γ-глобулини). Комплексирането с протеини е обратим процес. В допълнение, някои от хормоните в кръвта са свързани с корпускулите, по-специално еритроцитите.
Образуването на свързана форма на хормони е от голямо физиологично значение. Първо, той предпазва тялото от излишното натрупване в кръвта (и следователно въздействието върху тъканите) на свободни хормони. На второ място, асоциираната форма на хормона е неговият физиологичен резерв. Трето, свързването с протеини помага да се предпази хормонът от разграждане от ензими, т.е. удължава живота му. И накрая, комплексирането с протеини предотвратява филтрирането на малки молекулярни хормони през бъбречните гломерули и по този начин ограничава тези важни регулаторни процеси.
4.
Разпознаване на хормоналния сигнал.След попадане в периферните органи, хормоните, като правило, се освобождават от протеиновия компонент и, фиксирайки се върху определени рецептори на клетките, които възприемат дадения хормон (целеви клетки), извършват специфичното си действие. В процеса на периферното действие на хормоните се случват техните различни трансформации. В този случай е възможно образуването на нови хормонални продукти, често по-активни или произвеждащи различен биологичен ефект от първоначалния хормон. По този начин тиреоидният хормон тироксин може да се превърне в тъканите в трийодтиронин, по-активен хормон на същата жлеза. Андрогените (мъжките полови хормони) се превръщат в хипоталамуса в естрогени - женски полови хормони. Трансдукцията на хормонален сигнал в биологичен отговор е органично свързана с механизма на действие на този хормон.
Действието на всеки хормон върху прицелните клетки винаги започва с взаимодействието му с определени компоненти на клетката. Това явление се нарича прием на хормони, а клетъчните компоненти, които взаимодействат с хормона, се наричат \u200b\u200bрецептори. Хормоналните рецептори са киселинни олигопептиди с голямо молекулно тегло.
Структурата на рецепторната молекула се характеризира с асиметрия. Има три раздела от него:
свързващ хормон;
ефектор, предаващ хормонален сигнал към вътреклетъчните механизми, т.е. преобразуване на сигнал в биологичен отговор;
свързване на първата и втората секции.
- хипофизната жлеза синтезира / секретира растежен хормон (STH), пролактин, ACTH и др .;
- в надбъбречната жлеза има четири слоя клетки, всеки от които синтезира свой собствен хормон.
От гледна точка на гастроентеролога панкреасът е екзокринен орган, тъй като секретира панкреатични ензими; от гледна точка на ендокринолог, той е ендокринен орган, тъй като произвежда пакет от взаимозависими хормони (инсулин, глюкагон, соматостатин и др.).
Освен това някои хормони се произвеждат на няколко места:
- катехоламини - не само в мозъка на надбъбречната жлеза, но и в ганглиите на паравертебралния нерв;
- соматостатин - както в островчетата Лангерханс, така и в хипоталамуса.
Извън ендокринните жлези са открити микроскопични клъстери от клетки, които са специализирани в синтеза на биологично активни вещества с хормонални свойства:
- регулатори на секрецията на хормони на ендокринната жлеза:
- ядрата на хипоталамуса синтезират вещества, които регулират секрецията на хипофизни хормони (соматолиберин, АСТН-освобождаващ хормон и др.);
- натрупвания на клетки в чревната стена, които произвеждат инкретинови хормони;
- регулатори на функциите на органите:
Относително наскоро бяха открити биологично активни вещества лептин и адипонектин, синтезирани от мастна тъкан (адипоцити), които бяха класифицирани като хормони, тъй като имат системен регулаторен ефект - регулират апетита и енергийния метаболизъм.
И така, хормоните се произвеждат не само от ендокринната жлеза, в резултат на което това качество не може еднозначно да дефинира понятието „хормон“. В същото време в съвременната клинична ендокринология почти всички заболявания представляват едно или друго нарушение на функциите на ендокринната жлеза. В тази връзка дефиницията на хормона и свързаната с нея дефиниция на ендокринната жлеза в клиничната ендокринология все още е „класическа“.
По този начин можем да дадем следното, напълно пълно от клинична гледна точка, определение на хормона.
Хормон - биологично активно вещество, произведено от ендокринната жлеза, което има регулаторен ефект върху определени структури на тялото и метаболизма (използване на субстрати от кръвта, енергиен обмен и др.), което често се проявява чрез видими външно промени в тялото (например растеж) и / или промени в поведението ( например сексуален).
В тази класическа дефиниция термините ендокринна жлеза и хормон са взаимозависими. Следователно логиката на диагностичното търсене в клиничната ендокринология е очевидна - чрез изследване на кръвните хормони за диагностициране на заболявания на жлезите с вътрешна секреция.
Определение за ендокринната жлеза
Ендокринна жлеза - добре дефинирана макроанатомична структура, чиято основна функция е синтезът на биологично активни вещества, наречени хормони. В клиничната ендокринология се разграничават седем жлези с вътрешна секреция, функциите на които се оценяват чрез изследване на хормони, произвеждани от жлезата в кръвта. За оценка на неговите функции се използва не целият спектър на хормоните на жлезите, а строго ограничен набор от тях, с помощта на които се определя функцията на ендокринната жлеза. В допълнение към хормоните, техните метаболити могат да се използват за диагностициране на заболявания, които понякога се оказват по-надежден маркер на ендокринната болест от изследването на самите хормони. Така че, при диагностицирането на феохромоцитома, изследването на метанефриновите катехоламинови метаболити е по-надеждно от адреналина и норепинефрина.
Изследването на хормони за диагностика на ендокринни заболявания не винаги е оправдано. Най-поразителният пример е захарният диабет, при диагностицирането на който не се използва изследването на инсулина, въпреки че заболяването се причинява от дефицит на инсулин. Също така, изследването на окситоцин и вазопресин не се използва за диагностициране на тяхната недостатъчна или прекомерна секреция, а нарушението на техния синтез се определя от техните метаболитни ефекти.
Освен това при диагностицирането на ендокринни заболявания могат да се използват хормони, които не се синтезират от жлезите с вътрешна секреция, например инсулиноподобен растежен фактор I (IGF-I), който се образува в черния дроб под действието на STH. Използва се за диагностициране на акромегалия, причинена от тумор на хипофизата.
Синтезът на хормона от ендокринната жлеза може да бъде:
- единствената му функция (например, предния лоб на хипофизната жлеза);
- комбиниран с генерирането на зародишни клетки (например яйчници и тестиси);
- комбиниран с екзокринна секреция (например панкреаса);
- в съчетание с отлагането на хормони, синтезирани извън него.
Ендокринната жлеза е в състояние да синтезира:
- единственият рядък хормон (например паращитовидната жлеза);
- спектър на хормоните (обикновено):
- чрез специализирани клетъчни субструктури, по-специално в надбъбречните жлези, две клетъчни субструктури - кората и медулата - произвеждат съответно стероидни хормони и катехоламини;
- отделни клетки, обединени или не в изолирани комплекси, например в хипофизната жлеза, някои хормони синтезират отделни клетки, които не са комбинирани в отделни клетъчни образувания; в панкреаса инсулинът и глюкагонът се произвеждат от β- и α-клетки, обединени в островчетата Лангерханс.
Същността и функцията на хормоните
Хормоните са разделени на две основни групи.
Полипептиди или производни на аминокиселини (най-много):
- сложни полипептиди (LH, hCG);
- средно големи пептиди;
- малки пептиди;
- дипептиди (Т4 и ТЗ);
- производни на отделни аминокиселини (серотонин, хистамин).
Производните на холестерол са стероиди от два вида:
- с непокътнат стероиден пръстен (надбъбречни и гонадни стероиди);
- с несвързан пръстен B.
Има четири основни функции на хормоните в организма:
- възпроизвеждане;
- растеж и развитие;
- производство, оползотворяване и съхранение на енергия.
Отделен хормон, от една страна, може да има различни биологични ефекти върху различни органи, а в един орган, в различно време; от друга страна, някои биологични процеси са под интегрален контрол на няколко хормона.
Хормоните регулират функциите на следните цели:
- други жлези с вътрешна секреция (напр. хипофизно-надбъбречната жлеза);
- функционални системи;
- органи (напр. Т4 и сърдечна функция или Т4 и мозъчна функция);
- тъкани (като кортизол и кости).
Синтез, съхранение и секреция на хормони
Пептидните хормони се синтезират по същия механизъм като всички други протеини. Често първо се синтезира голяма прохормонна молекула, която след това се превръща в по-малък хормон. Например препапаратиреоиден хормон → пропатиреоиден хормон → паратиреоиден хормон. От друга страна, стероидите и катехоламините се синтезират от по-малки молекули.
Ендокринните органи не са уникално място за синтеза на хормони, но само в тях синтезът на хормони и неговата регулация се случват най-ефективно. Три основни характеристики отличават ендокринния орган от неендокринната тъкан, която синтезира всеки хормон:
- скоростта на синтез е много по-висока в ендокринния орган;
- жлезите с вътрешна секреция са снабдени с механизъм за транспортиране на хормона в кръвта, който по правило се регулира.
Скоростта на хормонална секреция от жлезата се определя от скоростта на нейния синтез, който може да се регулира от други хормони, тропични за тази жлеза. С изключение на Т4 и 1,25-дихидроксихолекалциферол, запасите от хормони в организма са много ограничени.
Стимулирането на хормоналната секреция е свързано с деполяризация на клетъчната мембрана и отваряне на калциевите канали, което води до навлизането на калция в клетката, където той се комбинира с калциево свързващ протеин.
Транспорт и елиминиране на хормони
Хормоните се елиминират от кръвта в резултат на метаболитните процеси, например, пептидните хормони се инактивират от протеолитични ензими. В черния дроб хормоните се комбинират с глюкуроновата киселина и се секретират с жлъчката, но в същото време те се реабсорбират частично, като се включват в така наречения ентерохепатален цикъл. Хормоните също се отделят с урината.
Малки молекули хормони (в частност Т 4) се свързват с кръвните протеини, което забавя елиминирането им от кръвта и поддържа малък пул свободен хормон в кръвта на необходимото ниво. Свързването с протеини също улеснява транспорта на мастноразтворими стероиди.
Хормонални рецептори
Хормоналните рецептори са клетъчни протеини, които свързват хормон.
Взаимодействието с хормона причинява конформационна промяна в рецептора, която активира специфична клетъчна ензимна система, която всъщност реализира характерния ефект на хормона. Когато хормонът се свърже с рецептора на клетъчната мембрана, в цитозола се появяват т. Нар. Втори посланици (първият е хормонът). В клетъчното ядро, хормон-рецепторен комплекс е свързан с дезоксирибонуклеинова киселина (ДНК) и регулира генната експресия. Максималният ефект на хормона обикновено се проявява, когато са свързани по-малко от 50% от рецепторите. Свободните рецептори, освободени от връзката с хормона, се връщат отново в цитозола или в клетъчната мембрана, където продължават да участват в взаимодействието на хормона и рецептора.
Стероидните хормони са липофилни, поради което те свободно дифузират през клетъчната мембрана и след това се свързват с белтъците на цитозолния рецептор.
Т3 се свързва с белтъците на ядрените рецептори, а ТЗ-рецепторният комплекс, когато се комбинира с ДНК, стимулира образуването на информационна РНК. Често стероидните и щитовидните хормони действат синергично, взаимно подсилвайки специфични ефекти (усилвайки генната експресия).
Броят на рецепторите на клетъчните мембрани и вътреклетъчните рецептори се променя и силата на връзката им с хормона също се променя. Клетките на миометриума и млечните жлези съдържат окситоцинови рецептори, чийто брой се увеличава под действието на естроген (регулиране нагоре) и намалява под действието на прогестерон (регулиране надолу). Миокардът съдържа норепинефринови рецептори (β 1), чийто брой и афинитетът към норепинефрин се увеличават под действието на тиреоидни хормони (Т 3 / Т 4).
Водоразтворимите хормони (моноамини, аминокиселини и пептиди) се свързват с рецепторите на мембраната, която е наситена с липиди, поради което не позволява на водоразтворимите хормони да дифузират свободно през мембраната. В хормоналната реакция на клетката водоразтворимите хормони се наричат \u200b\u200bпървите пратеници. В отговор на взаимодействието им с рецептора вътре в клетката се активират т. Нар. Втори посланици - сАМР, цикличен гуанозин монофосфат, инозитол трифосфат, калциеви йони, диацилглицерол и др. Калциевите йони служат като много важен втори посланик. Потокът на калциевите йони през клетъчната мембрана в цитозола се контролира чрез комуникация хормон-рецептор, нервни дразнители или модифициран от други втори посланици.
Концентрацията на хормоните в повечето случаи е 10 -10 mol / l. В този случай свързването на една молекула с мембранния рецептор води до образуването на 10 000 сАМР молекули в клетката и в това отношение сАМР действа като молекулярен усилвател на хормоналния сигнал (10 000 пъти!). Фосфодиестеразата унищожава сАМР, така че нейните инхибитори теофилин и кофеин действат синергично с хормоните, при които сАМР е вторият пратеник. cAMP стимулира катаболните процеси - липолиза, гликогенолиза (глюкагон), глюконеогенеза и кетогенеза, секреция на инсулин в β-клетките и панкреаса.
Резюме по темата
Ендокринни жлези и техните хормони
Ендокринни жлези
(от гръцки endon - вътре, crio - разпределям) или жлези с вътрешна секреция, са специализирани органи или групи клетки, чиято основна функция е да развиват и освобождават специфични биологично активни вещества във вътрешната среда на тялото. Ендокринните жлези нямат отделителни канали. Клетките им са плетени от богата мрежа от кръвоносни и лимфни съдове, а отпадъчните продукти се отделят директно в кръвта, лимфата и тъканната течност. Тази характеристика фундаментално отличава жлезите с вътрешна секреция от жлезите с външна секреция, които отделят секрета си чрез отделителните канали. Продуктите, произведени от жлезите с вътрешна секреция, се наричат хормони(Гръцки hormao - възбуждам, активирам). Терминът "вътрешна секреция" е предложен през 1885 г. от френския физиолог К. Бернар, а терминът "хормон" - от английските физиолози W. Beilis и E. Starling през 1902 г.
Следните характеристики са характерни за хормоните: Техният синтез и изолиране се извършва от специализирани клетки.
Хормоните се образуват в жлезистите ендокринни клетки, след което попадат във вътрешната среда, главно в кръвта и лимфата. Междинните продукти от синтеза или метаболизма на хормоните често имат биологична активност, но като правило те не са серкетирани .
Висока биологична активност.
Хормоните имат физиологичен ефект при много ниски концентрации. И така, концентрацията на женския полов хормон (естрадиол) в кръвта варира от 0,2 до 0,6 μg (10 -6 g) в 100 ml плазма. Съдържанието на растежен хормон в кръвта се измерва в още по-малки количества - нанограми (KG 9 g). Хипофизната жлеза реагира на пикограми (10 -12 g) хипоталамусни хормони, ангиотензин-Р-продукт на ендокринни бъбречни клетки - предизвиква чувство на жажда при фемтограми (10 -15 g). Освен хормоните, никакви други химически отпадъчни продукти не са ефективни в такива малки дози. Специфичност.Всеки хормон се характеризира със специфична, присъща само химическа структура, място на синтез и функция. В тази връзка дефицитът на който и да е хормон не може да бъде попълнен с други хормони или биологично активни вещества. Действие от разстояние.
Хормоните, като правило, се носят с кръв далеч от мястото на образуване, засягайки отдалечени органи и тъкани. По този начин те се различават от медиаторите и цитокините, действащи върху една клетка или група клетки на мястото на тяхното образуване.
Химичната структура на хормоните и тяхната трансформация в организма
Според своята химическа структура хормоните могат да бъдат разделени на 4 основни групи:
протеини и пептиди;
производни на аминокиселини;
стероиди;
простагландини.
Примерите за протеинови хормони включват инсулин, соматотропин (хормон на растежа), тропни хормони на предната хипофизна жлеза. Някои от тях (фолитропин, тиротропин, лутропин) са сложни протеини, други (инсулин, калцитонин и др.) Са прости протеини. Пептидната структура има глюкагон, вазопресин, окситоцин, хипоталамусни хормони. Производните на аминокиселини включват хормони на щитовидната жлеза - трийодтиронин, тироксин, както и адреналин и норепинефрин. Стероидните хормони се основават на ядрото на цикличния въглеводород циклопентанперхидрофенантрен. Тази група включва хормони на надбъбречната кора и половите жлези. Основните етапи на образуването и трансформацията на хормона могат да бъдат представени както следва:
биосинтеза на хормони;
секреция, т.е. изолация от ендокринна клетка;
пренос на кръв към периферните тъкани;
разпознаване на хормоналния сигнал от целевите клетки;
трансдукция (транслация) на хормонален сигнал в биологичен отговор;
потискане на хормоналния сигнал.
Органи и тъкани с клетъчна ендокринна функция
№
|
Органи и тъкани
|
Клетки
|
Хормони
|
1.
|
Плацента
|
Синцитиотрофобласти Цитотрофобласти
|
Хорион гонадотропин, пролактин Естриол, прогестерон
|
2.
|
Тимус
|
Тимоцити
|
Тимозин, тимопоетин
|
3.
|
Пъпка
|
ЮГА Интерстициум
Тръбички
|
Ренин Еритропоетин
Калцитриол
|
4.
|
Сърце
|
Предсърдни миоцити
|
Атриопептид, соматостатин, ангиотензин-Р
|
Органи, тъкани и клетки с ендокринна функция
№
|
Структури
|
Тъкани, клетки
|
Хормони
|
1.
|
Хипоталамус
|
Нервни клетки на структури с малка точка
|
Хипоталамусни неврохормони:
|
|
|
|
Заден хипоталамус Нервни клетки на големи клетъчни ядра на предния хипоталамус
|
а) либерини: кортиколиберин, тиролиберин, лулиберин, фолиберин, соматолиберин, пролактолиберин, меланолиберин; б) статини: пролактостатин, меланостатин, соматостатин
Вазопресин, окситоцин
|
2.
|
Хипофиза Аденохипофиза Неврохипофиза
|
Кортикотрофи Гонадотрофи
Тиротрофи
Соматотрофи
Лактотрофи
Питуицити
|
Кортикотропин, меланотропин Фолитропин, лутропин Тиротропин Растежен хормон Пролактин
Вазопресин, окситоцин
|
3.
|
Надбъбречни жлези Кортиково вещество
Мозъчна материя
|
Зона на гломерулите Зона на сноп Мрежа зона Хромафинови клетки
|
Минералокортикоиди Глюкокортикоиди Полови стероиди: андрогени, естрогени Андреналин, норепинефрин
|
4.
|
Щитовидна жлеза
|
Фоликуларни тироцити К клетки
|
Трийодтиронин, тетрайодтиронин Калцитонин, катакалцин, Ко-калцигенин
|
5.
|
Паращитовидните жлези
|
K-клетъчни основни клетки
|
Паратирин калцитонин
|
6.
|
Епифиза
|
Пинеоцити
|
Мелатонин
|
7.
|
Панкреас
|
Островчета Лангерханс: осини клетки R-клетки А-клетки
|
Глюкагон Инсулин
Соматостатин
|
8.
|
Полови жлези (тестиси, яйчници)
|
Leydig клетки Sertoli клетки Granulosa клетки Жълто тяло
|
Тестостерон естрогени, инхибин естрадиол, естрон, прогестерон Прогестерон
|
Биосинтеза на хормонипрограмирани в генетичния апарат на специализирани ендокринни клетки. Следователно, това зависи от структурата и експресията на гени, кодиращи синтеза на тези хормони, както и от ензимите, които регулират синтеза на хормони и пост-транслационните процеси. Липсата или дефектът на съответните гени води до ендокринопатия. Пример за това е нанизъм с генетичен дефект на хормона на растежа.
2. Секреция на хормони.Както е посочено, най-важната характеристика на всеки хормон е неговата секреция. Опакован във везикули или гранули, хормонът се транспортира към цитоплазматичната мембрана. Освобождаването на протеин-пептидни хормони и катехоламини от клетката се предшества от взаимодействието на цитоплазматичната мембрана и мембраната на секреторната гранула. След това се случва техният лизис и хормонът се освобождава от клетката. Този процес се активира от много фактори, медиатори, висока концентрация на калий, електрически стимули и др. Хормоналната секреция е акт, придружен от разход на енергия, поради което винаги е свързан с промени в системата АТФ - цАМФ. За секрецията е необходимо участието на калциеви йони, които активират протеините на микротубуларно-микрофиламентозната система, насърчавайки взаимодействието на хормонални гранули с тях и влияят върху образуването на цАМФ. Следователно намаляването на съдържанието на калциеви йони в извънклетъчната среда и навлизането му в ендокринната клетка неизбежно води до намаляване на секреторната активност на тази клетка. 3.
Транспорт на хормони.Секретираният хормон навлиза във вътрешната среда на тялото, главно в кръвта, и се транспортира по-нататък. Повечето хормони образуват сложни съединения с плазмени протеини в кръвта. Някои от тези протеини са специфични транспортни протеини (например транскортин, който свързва хормоните на надбъбречната кора), а някои са неспецифични (например γ-глобулини). Комплексирането с протеини е обратим процес. Освен това някои от хормоните в кръвта са свързани с корпускулите, по-специално еритроцитите. Образуването на свързана форма на хормони е от голямо физиологично значение. Първо, той предпазва тялото от излишното натрупване в кръвта (и следователно въздействието върху тъканите) на свободни хормони. На второ място, асоциираната форма на хормона е неговият физиологичен резерв. Трето, свързването с протеини помага да се предпази хормонът от разграждане от ензими, т.е. удължава живота му. И накрая, комплексирането с протеини предотвратява филтрирането на малки молекулни хормони през бъбречните гломерули и по този начин ограничава тези важни регулаторни процеси.
4.
Разпознаване на хормоналния сигнал.След попадане в периферните органи, хормоните, като правило, се освобождават от протеиновия компонент и, фиксирайки се върху определени рецептори на клетките, които възприемат дадения хормон (целеви клетки), извършват специфичното си действие. В процеса на периферното действие на хормоните се случват техните различни трансформации. В този случай е възможно образуването на нови хормонални продукти, често по-активни или произвеждащи различен биологичен ефект от първоначалния хормон. По този начин тиреоидният хормон тироксин може да се превърне в тъканите в трийодтиронин, по-активен хормон на същата жлеза. Андрогените (мъжките полови хормони) се превръщат в хипоталамуса в естрогени - женски полови хормони. Трансдукцията на хормонален сигнал в биологичен отговор е органично свързана с механизма на действие на този хормон. Действието на всеки хормон върху прицелните клетки винаги започва с взаимодействието му с определени компоненти на клетката. Това явление се нарича прием на хормони, а клетъчните компоненти, които взаимодействат с хормона, се наричат \u200b\u200bрецептори. Хормоналните рецептори са киселинни олигопептиди с голямо молекулно тегло.
Структурата на рецепторната молекула се характеризира с асиметрия. Има три раздела от него:
свързващ хормон;
ефектор, предаващ хормонален сигнал към вътреклетъчните механизми, т.е. преобразуване на сигнал в биологичен отговор;
свързване на първата и втората секции.
Рецепторите са разположени или вътреклетъчно, или на повърхността на цитоплазмената мембрана. Вътреклетъчните рецептори свързват стероидни и щитовидни хормони, мембранните рецептори свързват други хормони. Също така се смята, че функцията за разпознаване на специфичен хормонален сигнал във всички клетки за всички хормони се осъществява от мембранния рецептор и след като хормонът се свърже със съответния му рецептор, по-нататъшната роля на хормонално-рецепторния комплекс за пептидни и стероидни хормони е различна. В пептида, протеиновите хормони и катехоламините хормон-рецепторният комплекс води до активиране на мембранните ензими и образуването на различни вторични медиатори (пратеници) на хормоналния регулаторен ефект, които реализират своето действие в цитоплазмата, органоидите и клетъчното ядро.
Известни са четири системи на вторични посредници:
аденилат циклаза - цикличен аденозин монофосфат (сАМР);
гуанилат циклаза - цикличен гуанозин монофосфат (cGMP);
фосфолипаза С - инозитол трифосфат (ITP);
йонизиран калций.
4.1.
Системата "аденилат циклаза - сАМР".
Мембраните ензимът аденилат циклаза могат да бъдат в две форми - активирана и неактивирана. Активирането на аденилат циклазата се осъществява под въздействието на хормон-рецепторен комплекс, чието образуване води до свързване на гуанил нуклеотид (GTP) със специален регулаторен стимулиращ протеин (GS-протеин), след което GS-протеинът причинява свързването на магнезий към аденилат циклазата и неговото активиране. Ето как хормоните, които активират аденилатциклазата - глюкагон, тиротропин, паратирин, вазопресин, гонадотропин и др. Някои хормони, напротив, потискат аденилат циклазата (соматостатин, ангиотензин-Р и др.). Под въздействието на аденилатциклаза от АТФ се синтезира сАМР, който активира протеинкинази в цитоплазмата на клетката, които осигуряват фосфорилиране на множество вътреклетъчни протеини. Това променя мембранната пропускливост, т.е. причинява метаболитни и съответно функционални промени, характерни за хормона. Вътреклетъчните ефекти на сАМР се проявяват и във влиянието върху процесите на пролиферация, диференциация, върху наличието на мембранни рецепторни протеини за хормонални молекули.
Гуанилат циклаза - cGMP система.
Активирането на мембранната гуанилатциклаза се случва не под прякото влияние на хормоно-рецепторния комплекс, а индиректно чрез йонизирани калциеви и оксидантни мембранни системи. Ето как предсърдният натриуретичен хормон - атриопептидът, тъканният хормон на съдовата стена, осъзнава своите ефекти. В повечето тъкани биохимичните и физиологичните ефекти на cAMP и cGMP са противоположни. Примерите включват стимулация на сърдечни контракции под въздействието на cAMP и инхибиране на техния cGMP, стимулиране на чревни контракции на гладката мускулатура с cGMP и потискане на cAMP.
4.3 Система "фосфолипаза С - йонозитол трифосфат".
Хормонно-рецепторният комплекс с участието на регулаторния протеин G води до активиране на мембранния ензим фосфолипаза С, което причинява хидролиза на мембранните фосфолипиди с образуването на два вторични пратеника - инозитол фосфат и диацилглицерол. Йозитол трифосфатът причинява освобождаването на йонизиран калций от вътреклетъчните запаси. Последният, свързвайки се с протеина калмодулин, активира протеинкинази и фосфорилиране на вътреклетъчни структурни протеини и ензими. Диацилглицеролът също активира и завършва процеса на фосфорилиране на други протеини, като същевременно реализира втория път на хормоналния ефект: чрез образуването на арахидонова киселина, която е източник на вещества, мощни по отношение на метаболитните и физиологични ефекти - простагландини и левкотриени. Ефектите на адреналин, вазопресин, ангиотензин-Р, соматостатин, окситоцин и някои други хормони се реализират чрез разглежданите системи на вторични медиатори.
4.4. Система "калций - калмодулин".
Йонизираният калций попада в клетката след образуването на хормон-рецепторен комплекс или от извънклетъчната среда поради активирането на бавни калциеви канали (например в миокарда), или от вътреклетъчните запаси под въздействието на гореописаните вътреклетъчни процеси. В цитоплазмата на немускулните клетки калцият се свързва със специален протеин - калмодулин, а в мускулните клетки ролята на калмодулина се играе от тропонин С. Калмодулинът, свързан с калция, активира множество протеинкинази, които осигуряват фосфорилиране на протеините. Краткосрочното увеличаване на количеството калций в клетката и свързването му с калмодулин е стимул за множество физиологични процеси - мускулна контракция, секреция на хормони и освобождаване на медиатори, синтез на ДНК, промени в клетъчната подвижност, ензимна активност и транспорт на вещества през мембраните. В стероидните хормони мембранният рецептор осигурява специфично разпознаване на хормона и прехвърлянето му в клетката. В цитоплазмата има специален цитоплазмен протеинов рецептор, с който се свързва хормонът. Тази връзка с рецепторния протеин е необходима за навлизането на стероидния хормон в ядрото, където той взаимодейства с третия ядрен рецептор, свързването на комплекса "хормон-ядрен рецептор" с хроматиновия акцептор, специфичен киселинен протеин и ДНК, което води до активиране на транскрипцията на иРНК, синтез на транспорт и рибозомна РНК, транспорт на иРНК в цитоплазмата, транслация на иРНК със синтеза на протеини и ензими в рибозомите. Всички тези явления изискват дългосрочно присъствие на хормон-рецепторния комплекс в ядрото. Ефектите от стероидните хормони обаче се проявяват не само след няколко часа, някои от тях се появяват бързо, в рамките на няколко минути. Това са ефекти като повишаване на мембранната пропускливост, увеличаване на транспорта на глюкоза и аминокиселини, освобождаване на лизозомни ензими, промени в енергията на митохондриите и, в допълнение, увеличаване на сАМР и йонизиран калций. По този начин възгледът е напълно оправдан, привържениците на който вярват, че мембранният рецептор за стероидни хормони не само изпълнява функцията на „разпознаване“ на хормоналната молекула, но подобно на рецепторите за пептидни хормони активира системата на вторичните пратеници в клетката. Пептидните хормони също имат способността да влияят селективно на генната транскрипция в клетъчното ядро. Този ефект може да се реализира не само от клетъчната повърхност поради вторични медиатори, но и чрез притока на хормони в клетката поради интернетизацияхормон-рецепторен комплекс (поради ендоцитоза).
Феноменът на интернализация на хормонално-рецепторните комплекси и по този начин намаляване на броя на хормоналните рецептори в клетъчната мембрана дава възможност да се разбере механизмът на намаляване на чувствителността на ефектора с прекомерно количество хормонални молекули или явлението десенсибилизацияефектор. Това явление всъщност представлява отрицателна регулаторна обратна връзка на ниво ефектор. Обратното явление е сенсибилизация,или увеличаване на чувствителността към хормони, може да се дължи на увеличаване на броя на свободните рецепторни места на мембраната, както поради намаляване на интернализацията, така и в резултат на „плаването“ на активните свързващи места на рецепторите. По този начин хормоните предават информационни сигнали на клетката и самата клетка е в състояние да регулира степента на възприемане на хормоналния контрол. 5. Предаване на хормонален сигнал в биологичен отговор.Има 5 вида хормонално действие върху прицелната тъкан: метаболитно, морфогенетично, кинетично, коригиращо, реактогенно. Метаболитно действие.
Метаболитните промени, причинени от хормони, са в основата на промените във функциите на клетките, тъканите и органите. Морфогенетично действие -
влиянието на хормоните върху процесите на образуване, диференциация и растеж на структурни елементи. Примерите включват ефекта на хормона на растежа върху растежа на тялото и вътрешните органи, ефекта на половите хормони върху развитието на вторични полови белези. Кинетично действие
- способността на хормоните да задействат активността на ефектора, да включва изпълнението на определена функция. Например, окситоцинът предизвиква свиване на мускулите на матката, адреналинът задейства разграждането на гликогена в черния дроб и освобождаването на глюкоза в кръвта, вазопресинът включва механизма на реабсорбция на вода в събирателните канали на нефроните. Коригиращи действия
- промени в дейността на органите или процеси, които се случват при липса на хормон. Примери за коригиращ ефект на хормоните са ефектът на адреналина върху сърдечната честота, активиране на окислителните процеси от тироксин и намаляване на реабсорбцията на калиеви йони в бъбреците под въздействието на алдостерон. Един вид коригиращо действие е нормализиращият ефект на хормоните, когато тяхното влияние е насочено към възстановяване на изменен или дори нарушен процес. Например, при първоначалното разпространение на анаболните процеси на протеиновия метаболизъм, глюкокортикоидите предизвикват катаболен ефект, но ако първоначално преобладава разграждането на протеини, глюкокортикоидите стимулират техния синтез. Реактогенно действие
- способността на хормона да променя реактивността на тъканта към действието на същия хормон, други хормони или медиатори на нервни импулси. Например, хормоните на щитовидната жлеза усилват ефекта на катехоламините, регулиращите калция хормони намаляват чувствителността на дисталния нефрон към действието на вазопресина. Вариация на реактогенното действие на хормоните е разрешителното действие, което предполага способността на един хормон да даде възможност за реализиране на ефекта на друг хормон. Така например, глюкокортикоидите имат разрешителен ефект по отношение на катехоламините (за реализиране на ефектите на адреналина е необходимо наличието на малки количества кортизол); инсулинът има пермисивен ефект върху соматотропин и др. 6. Потискане на хормоналния сигнал.Този механизъм действа за нормализиране на хормоналния сигнал (ако се забави, ще има опасност от прекомерно напрежение в много функции). Съществуват множество механизми за потискане на хормоналния сигнал. На първо място, това е намаляване на биосинтетичната и секреторна активност на самите клетки, произвеждащи хормони. Заедно с това се случва и разрушаването (катаболизма) на някои от секретираните хормони. В резултат значителна част от хормоните изчезват от кръвта, след като са били освободени от гранулите. Чернодробните ензими инактивират хормоните чрез дезаминиране и метилиране. В процеса на инактивация както хормоните, така и продуктите от техния катаболизъм се канюгират в черния дроб с глюкуронова и сярна киселини, което увеличава тяхната разтворимост във вода и улеснява последващата екскреция през бъбреците. Част от свободни и конюгирани хормони влиза в жлъчката и се екскретира от тялото през червата. Потискането на хормоналния сигнал се улеснява от десенсибилизацията на тъканите, описана по-горе с дългосрочно повишено ниво на хормона в кръвта. В някои случаи се образуват антитела към хормони и други инхибитори, които инхибират биологичния ефект.
Намаляване на активността на хормоналния сигнал се постига и в резултат на секрецията на контрарегулаторни хормони, т.е. хормони с противоположно действие. Например излишъкът от инсулин причинява хипогликемия, която от своя страна индуцира секрецията на контраинсуларни хормони. Друг важен физиологичен механизъм за потушаване на хормоналния сигнал е инхибирането на секрецията съгласно принципа на обратната връзка.
Съвременна теория на регулаторните пептиди
Регулаторни пептиди- биологично активни вещества, синтезирани от клетки на тялото от различен произход и участващи в регулирането на различни функции. Сред тях се различават невропептидите, които се секретират от нервните клетки и участват в изпълнението на функциите на нервната система. Освен това те се намират извън централната нервна система в редица ендокринни жлези, както и в други органи и тъкани.
В онтогенезата регулаторните пептиди се появяват много по-рано от „класическите“ хормони, т.е. преди изолирането на специализирани жлези с вътрешна секреция. Това ни позволява да приемем, че отделното образуване на посочените групи вещества е програмирано в генома и следователно те са независими.
Източници на регулаторни пептиди са единични клетки, продуциращи хормони, понякога образуващи малки клъстери. Тези клетки се считат за начална форма на ендокринни образувания. Те включват невросекреторни клетки на хипоталамуса, невроендокринни (хромафинови) клетки на надбъбречните жлези и параганглия, клетки на лигавицата на стомашно-чревната система, пинеалоцити на епифизната жлеза. Установено е, че тези клетки са способни да декарбоксилират ароматни киселини-прекурсори на невроамини, което дава възможност да се комбинират в една система (Pearse, 1976), наречена „APUD-система“ (от първите букви на английските думи Amine Precursor Uptake and Decarboxylating system - аминни предшественици). В секреторните елементи на стомашно-чревния тракт първоначално са открити голям брой пептиди (вазоактивен чревен пептид - VIP, холецистокинин, гастрин, глюкагон). Други (вещество Р, невротензин, енкефалини, соматостатин) първоначално са открити в нервната тъкан. Трябва да се отбележи, че в стомашно-чревния тракт някои пептиди (гастрин, холецистокинин, VIP и някои други) присъстват в нервите, както и в ендокринните клетки.
Съществуването на тази невродифузна ендокринна система се обяснява с миграцията на клетки от един източник - нервния гребен; те се включват в централната нервна система и тъканите на различни органи, където се трансформират в клетки, подобни на централната нервна система, секретиращи невроамини (невротрансмитери) и пептидни хормони. Това обяснява наличието на невропептиди в червата и панкреаса, клетките на Кулчицки в бронхите, а също така прави ясно появата на хормонално активни тумори на белите дробове, червата и панкреаса. Апудоцитите също се намират в бъбреците, сърцето, лимфните възли, костния мозък, епифизата и плацентата.
Основните групи регулаторни пептиди (според Кригер)
Най-често срещаната класификация на регулаторните пептиди включва следните групи:
хипоталамус освобождаващи хормони;
неврохипофизарни хормони;
хипофизни пептиди (ACTH, MSH, STH, TSH, пролактин, LH, FSH, (3-ендорфин, липотропини);
стомашно-чревни пептиди;
други пептиди (ангиотензин, калцитонин, невропептид V).
За редица пептиди бяха установени локализацията на съдържащите клетки и разпределението на влакната. Описани са няколко пептидергични системи на мозъка, които са разделени на два основни типа.
Дълги проекционни системи,
чиито влакна достигат до отдалечени области на мозъка. Например телата на невроните от семейство проопиомеланокортин са разположени в дъгообразното ядро \u200b\u200bна хипоталамуса и техните влакна достигат до амигдалата и околокръвното сиво вещество на средния мозък.
Системи за къса проекция:
телата на невроните често са разположени в много области на мозъка и имат локално разпределение на процесите (вещество Р, енкефалини, холецистокинин, соматостатин).
Много пептиди присъстват в периферните нерви. Например вещество P, VIP, енкефалини, холецистокинин, соматостатин се намират във блуждаещите, целиакия и седалищните нерви. Медулата на надбъбречната жлеза съдържа големи количества препроенкефалин А (метенкефалин). Показано е съществуването на невропептиди и невротрансмитери в същия неврон: серотонин е открит в невроните на продълговатия мозък заедно с вещество Р, допамин, заедно с холецистокинин, в неврони на средния мозък, ацетилхолин и VIP, в автономните ганглии. За функционалното значение на това съжителство може да се съди по следните фактори. Под въздействието на VIP при физиологични концентрации се наблюдава подчертано повишаване на чувствителността на мускариновите рецептори към ацетилхолин в подмандибуларната жлеза на котките, а антисерумът срещу VIP частично блокира вазодилатацията, причинена от стимулация на парасимпатиковите нерви.
Синтез на регулаторни пептиди.
Характерна особеност на пептидния синтез е образуването им чрез фрагментиране на голяма молекула-предшественик, т.е. в резултат на така нареченото пост-транслационно протеолитично разцепване - обработка. Синтезът на предшественика се случва в рибозомите, което се потвърждава от наличието на пратеник РНК, кодиращ пептида, и пост-транслационни ензимни модификации с освобождаването на активни пептиди - в апарата на Голджи. Тези пептиди достигат до нервните окончания чрез аксонален транспорт.
Активните пептиди, произхождащи от един предшественик, образуват неговото семейство. Описани са следните семейства пептиди.
Семейството на проопиомеланокортин (POMC).Телата на невроните, в които се намира този голям протеин (286 аминокиселинни остатъка) са локализирани в дъгообразното ядро \u200b\u200bна хипоталамуса. В зависимост от набора от ензими от POMC се образуват: в предния лоб на хипофизната жлеза - главно ACTH, (3-липотропин, P-ендорфин, в междинния - c-меланостимулиращ хормон и P-ендорфин. Това са ензимите катепсин В, трипсин, карбоксипептидаза, аминопептидаза, местата на тяхното нападение са сдвоени аминокиселинни остатъци.
Семейство Cerulein:гастрин, холецистокинин.
VIP семейство: секретин, глюкагон.
Семейство аргинин-вазопресин:вазопресин, окситоцин.
Освен това беше установено, че мет-енкефалинът и лев-енкефалинът имат прекурсори под формата на препроенкефалин А и препроенкефалин В, съответно. Протеолизата в този случай не е инактивиране, а трансформация на активността. Механизмът на действие на невропептидите:
Характерна особеност на регулаторните пептиди е полифункционалността (по отношение на механизма и естеството на ефектите) и образуването на регулаторни вериги (каскади). По принцип механизмите на действие на пептидите могат да бъдат разделени на две групи: синаптични и екстрасинаптични. 1.
Синаптични механизми на действие на пептидитеможе да се изрази в невротрансмитер или невромодулаторна функция. Невротрансмитер (Peyrotransmitter)- вещество, което се освобождава от пресинаптичния терминал и действа върху следващата - постсинаптична мембрана, т.е. изпълнява трансферна функция. Установено е, че някои пептиди изпълняват тази функция чрез пептидергични рецептори, присъстващи на невроните (техните тела или терминали). По този начин хипоталамусният освобождаващ хормон на лутеинизиращия хормон (лулиберин) в синаптичните ганглии на жабата се освобождава при стимулиране на нерва чрез калциево-зависим процес и причинява късен бавен възбуден постсинаптичен потенциал.
За разлика от "класическите" невротрансмитери (норепинефрин, допамин, серотонин, ацетилхолин), пептидите, които изпълняват трансферната функция, се характеризират с висок рецепторен афинитет (който може да осигури по-далечен ефект) и продължително (десетки секунди) действие поради липсата на ензимни инактивиращи системи и обратен депозит.
Невромодулатор,за разлика от невротрансмитера, той не предизвиква независим физиологичен ефект в постсинаптичната мембрана, но модифицира реакцията на клетката към невротрансмитера. По този начин невромодулацията не е трансферна функция, а регулаторна функция, която може да бъде изпълнена както на пост-, така и на пресинаптично ниво. Видове невромодулация:
контрол на освобождаването на невротрансмитер от терминали;
регулиране на невротрансмитерната циркулация;
модификация на ефекта на "класическия" невротрансмитер.
2.
Екстрасинаптично действие на пептидитереализиран по няколко начина. А. Паракринно действие(паракриния) - извършва се в зоните на междуклетъчен контакт. Например, соматостатинът, секретиран от А-клетките на островната тъкан на панкреаса, изпълнява паракринна функция в контрола на секрецията на инсулин и глюкагон (съответно от 3- и α-клетки и калцитонин - в контрола на секрецията на йодсъдържащи хормони от щитовидната жлеза. Б. Невроендокринно действие- осъществява се чрез освобождаването на пептида в кръвния поток и въздействието му върху ефекторната клетка. Примери за това са соматостатин и други хипоталамусни фактори, секретирани в медиалната еминенция от някои терминали в порталната циркулация и контролиращи секрецията на хипофизни хормони. Б. Ендокринно действие.В този случай пептидите се освобождават в общия кръвен поток и действат като отдалечени регулатори. Този механизъм включва компоненти, необходими за „класическите“ ендокринни функции - транспортни протеини и рецептори целеви клетки. Установено е, че като носители на стабилизатори се използват: неврофизини - за вазопресин и окситоцин, някои албумини и плазмени глобулини - за холецистокинин и гастрин. Що се отнася до приемането, е установено съществуването на отделни рецептори за опиоидни пептиди, вазопресин, VIL. Като вторични пратеници могат да се използват циклични нуклеотиди, продукти на хидролиза на фосфоинозитиди, калций и калмодулин с последващо активиране на протеин киназа и контрол на фосфорилирането на транслационни и транскрипционни регулаторни протеини. В допълнение е описан механизмът на интернализация, когато регулаторен пептид, заедно с рецептор, навлиза в клетката чрез механизъм, подобен на пиноцитозата, и сигнал се предава на невронния геном. Регулаторните пептиди се характеризират с образуването на сложни вериги или каскади в резултат на факта, че метаболитите, образувани от основния пептид, също са функционално активни. Това обяснява продължителността на ефектите от краткотрайните пептиди.
Функции на регулаторните пептиди: 1. Болка.Редица пептиди влияят върху образуването на болка като сложно психофизиологично състояние на тялото, включително самото усещане за болка, както и емоционални, волеви, двигателни и вегетативни компоненти. В този случай пептидите са включени както в ноцицептивната, така и в антиноцицептивната система. По този начин веществото P, соматостатин, VIP, холецистокинин и ангиотензин се намират в първичните сензорни неврони, а веществото P е невротрансмитер, секретиран от определени класове аферентни неврони. В същото време енкефалини, вазопресин, ангиотензин и свързаните с тях опиоидни пептиди се намират в низходящия супраспинален път, водещ до задните рога на гръбначния мозък и оказващ инхибиторен ефект върху ноцицептивните пътища (аналгетичен ефект).
2. Памет, учене, поведение.Има доказателства, че липсват фрагменти от ACTH (ACTH 4-7 и ACTH 4-10) хормонални ефекти, и ss-меланостимулиращият хормон подобряват краткосрочната памет, а вазопресинът участва в формирането на дългосрочната памет. Инжектирането на антитела срещу вазопресин в мозъчните вентрикули в рамките на един час след тренировката предизвиква забрава. Освен това ACTH 4-10 подобрява вниманието.
Установено е влиянието на редица пептиди върху хранителното поведение. Примерите включват повишаване на хранителната мотивация под действието на опиоидни пептиди и затихване под действието на холецистокинин, калцитонин и кортиколиберин.
Опиоидните пептиди имат значителен ефект върху емоционалните реакции, като са ендогенни еуфоригени.
VIP има хипнотичен, хипотензивен и бронходилататорен ефект. Тиреолиберинът има психотонен ефект. Лулиберин, освен че изпълнява командна функция (стимулиране на гонадотропите на предната хипофизна жлеза), регулира сексуалното и родителското поведение.
3. Вегетативни функции.Различни пептиди участват в контрола на нивото кръвно налягане... Това е ренин-ангиотензиновата система, всички компоненти на която присъстват в мозъка, опиоидни пептиди, VIP, калцитонин, атриопептид, които имат силен натриуретичен ефект.
Описани са промени в терморегулацията под действието на някои пептиди. И така, интрацентрално приложение на тиролиберин и Rβ-ендорфинът причинява хипертермия, докато прилагането на ACTH и α-MSH индуцира хипотермия.
4. Стрес.Забележително е, че редица невропептиди (опиоидни пептиди, пролактин, епифизни пептиди) са класифицирани като антистрес система, тъй като ограничават развитието на стресови реакции. По този начин, в експерименти с различни модели, беше показано, че опиоидните пептиди ограничават активирането на симпатиковото разделение на нервната система и всички връзки на хипоталамо-хипофизарно-надбъбречната система, предотвратявайки изчерпването на тези системи, както и нежеланите последици от излишните глюкокортикоиди (потискане на възпалителната реакция и тимо-лимфната система, появата на язви стомашно-чревния тракт и други) - Антихипоталамичните фактори на епифизата инхибират образуването на либерини и секрецията на хормони на предната хипофизна жлеза. Намаленото активиране на хипоталамуса ограничава хиперсекрецията на вазопресин, което има увреждащ ефект върху миокарда.
5. Влияние върху имунната система.Установени са двупосочни връзки между системата на регулаторните пептиди и имунната система. От една страна, способността на много пептиди да модулират имунните отговори е достатъчно проучена. Потискане на синтеза на имуноглобулини под действието на (3-ендорфин, енкефалини, ACTH и кортизол; инхибиране на секрецията на интерлевкин -1
(I Л -1)
и развитието на треска под въздействието на а-меланоцит-стимулиращ хормон. Установено е, че вазоактивният чревен пептид (VIL) инхибира всички функции на лимфоцитите и тяхното освобождаване от лимфните възли, което се разглежда като нова форма на имуномодулация. В същото време редица пептиди имат стимулиращ ефект върху имунната система, причинявайки увеличаване на синтеза на имуноглобулини и γ-интерферон (| 3-ендорфин, тиреоид-стимулиращ хормон), увеличаване на активността на естествените клетки убийци (R-ендорфин, енкефалини), увеличаване на пролиферацията на лимфоцити и освобождаването на лимфокини (вещество Р, пролактин, растежен хормон), увеличаване на производството на супероксидни аниони (растежен хормон). Описани са рецептори на лимфоцити за редица хормони. От друга страна, имунните медиатори влияят върху метаболизма и секрецията на хипоталамусните невротрансмитери и освобождаващи хормони. По този начин регулаторният левкопептид IL -1
е в състояние да навлезе в мозъка през зони с повишена пропускливост на кръвно-мозъчната бариера и да стимулира секрецията на кортикотропин-освобождаващ хормон (в присъствието на простагландин), последвано от стимулиране на освобождаването на ACTH и кортизол, които инхибират образуването на IL -1
и имунния отговор.
Едновременно с това чрез освобождаването на соматостатин IL -1
инхибира секрецията на TSH и хормона на растежа. По този начин имунопептидът действа като спусък, който чрез затваряне на механизма за обратна връзка предотвратява излишността на имунния отговор.
Според съвременните концепции пълният регулаторен кръг между невроендокринните и имунните механизми включва и пептиди, общи за двете системи. По-специално е показана способността на хипоталамусните неврони да секретират IL-1. Изолиран е генът, отговорен за неговото производство, чиято експресия се индуцира от бактериални антигени и кортикотропин. Описани са невронни пътища в човешкия и плъхов медиобазален хипоталамус, съдържащ IL-1 и IL-6, както и хипофизни клетки, секретиращи тези пептиди.
По този начин имунните медиатори могат да регулират функциите на предната хипофизна жлеза чрез:
ендокринен механизъм (лимфокини на активирани лимфоцити, циркулиращи в кръвта);
невроендокринни ефекти, реализирани от интерлевкини на хипоталамуса чрез тубероинфундибуларната портална система;
паракринен контрол в самата хипофизна жлеза.
От друга страна, резултатите от имунохимични и молекулярни изследвания показват, че имунокомпетентните клетки секретират много пептиди и хормони, свързани с ендокринната и невроналната активност: лимфоцитите и макрофагите синтезират АСТН; лимфоцити - хормон на растежа, пролактин, TSH, енкефалини; мононуклеарни лимфоцити и мастоцити - VIP, соматостатин; тимусни клетки - аргинин, вазопресин, окситоцин, неврофизин. В този случай хипофизните хормони, секретирани от лимфоцитите, се регулират от същите фактори като хипофизната жлеза. Например, секрецията на ACTH от лимфоцитите се инхибира от глюкокортикоиди и се стимулира от освобождаващия кортикотропин хормон. Предложена е концепция, според която освобождаването на изброените хормони от лимфоцитите осигурява автокринна и паракринна регулация на локалния имунен отговор. По този начин функциите на трите основни регулаторни системи - нервна, ендокринна и имунна - са интегрирани в сложни регулаторни кръгове, които функционират според принципа на обратната връзка. В същото време периферните лимфоцити, съгласно концепцията на Д. Блалок (Blalock, 1989), осигуряват сензорен механизъм, чрез който се разпознават некогнитивни стимули (чужди вещества) и се мобилизират невроендокринните адаптивни реакции.
Участие на регулаторни пептиди в развитието на патологията.
Тъй като пептидните хормони представляват полифункционална система, участваща в регулирането на много функции в тялото, вероятно е те да участват в патогенезата на различни заболявания. По този начин е установено нарушение на концентрациите на мозъчни пептиди при дегенеративни неврологични заболявания с неизвестна етиология: болест на Алцхаймер (намаляване на концентрацията на соматостатин в кората на главния мозък) и Хънтингтън (намаляване на концентрацията на холецистокинин, вещество Р и енкефалини, увеличаване на съдържанието на соматостатин в броя на базалните ганглии, както и на базалните рецептори свързване на холецистокинин в тези структури и в мозъчната кора). Дали тези промени са първични или се явяват като последица от развитието на болести, предстои да разберем.
Откриването на опиоидни пептиди и разпределението на техните рецептори в различни мозъчни структури, по-специално в лимбичната система, насочи вниманието към оценката на тяхното значение в патогенезата на психичните разстройства. Предлага се хипотезата за съществуването на опиоиден дефицит при пациенти с шизофрения, по-специално невъзможността за образуване на γ-ендорфин, който има невролептичен ефект. Установено е повишаване на концентрацията на атриопептид по време на стагнация в кръвоносната система, което вероятно е механизъм за компенсиране на нарушения на натриевия метаболизъм (неговото забавяне).
Изследването на олигопептидните хормони като регулаторна система доведе до изолиране на специална група заболявания, причинени от нейната патология - апудопатии.
Апудопатия- заболявания, свързани с нарушение на структурата и функцията на апудоцитите и изразени в определени клинични синдроми. Има първични апудопатии, причинени от патологията на самите апудоцити, и вторични апудопатии, възникващи като реакция на апудоцити към нарушение на хомеостазата на организма, причинено от заболяване, чиято патогенеза не е свързана предимно с патологията на системата APUD (при инфекциозни заболявания, растеж на тумори, заболявания на нервната система и др.) ). Първичните апудопатии могат да се проявят в хиперфункция, хипофункция, дисфункция и в образуването на апудоми - тумори от клетки на системата APUD. Примери за това са следните абдудоми.
Гастрином- апудом от клетки, които произвеждат гастрин, за който е известно, че стимулира секрецията голям брой стомашен сок с висока киселинност и храносмилателна сила. Следователно, клинично гастриномът се проявява чрез развитието на улцерогенен синдром на Zollinger Ellison. Кортикотропином- апудом, развиващ се от апудобласти на стомашно-чревния тракт и проявен чрез ектопична хиперпродукция на АСТН и развитие на синдрома на Иценко-Кушинг. Випома- тумор от клетки, секретиращи вазоактивен чревен пептид. Локализиран в дванадесетопръстника или панкреаса. Проявява се от развитието на водниста диария и дехидратация, както и от нарушение на метаболизма на електролитите. Соматостатином- тумор от чревни клетки или островна тъкан на панкреаса, произвеждащ соматостатин. Соматостатиномът обикновено се развива като тумор на панкреатичните D клетки, които секретират соматостатин. Характеризира се с клиничен синдром, включващ захарен диабет, жлъчнокаменна болест, хипохлорхидрия, стеаторея и анемия. Диагностицира се чрез повишаване на концентрацията на соматостатин в кръвната плазма. Приложение на регулаторните пептиди в медицината
Въз основа на регулаторни пептиди, някои лекарства... По този начин олигопептидите (къси пептиди) на N-крайния фрагмент на ACTH и MSH се използват за коригиране на вниманието и запаметяването, вазопресин - за подобряване на паметта при травматични и други амнезии. Домашното лекарство даларгин (аналог на леенкефалин) се използва широко в медицинската практика. Търговското освобождаване на сурфагон (аналог на лулиберин), предназначен за корекция на нарушения на репродуктивната система, започна. |