Žlijezde i njihovi hormoni u tijelu
U ljudskom tijelu, pored glavnih funkcionalnih organa, postoje i takozvane žlijezde, odnosno organi dizajnirani za sintezu i izlučivanje aktivnih supstanci zvane tajne. Žlijezde su često mikroskopske, a samo nekoliko je relativno velikih. Zanimljivo je da se žlijezde koje imaju izvodne kanale i izlučuju tajnu na sluznicu ili površinu tijela (znoj, pljuvačka, mlijeko i druge) zovu se egzokrine, a nemaju kanale i oslobađaju visoko aktivne tvari (hormone) direktno u krvožilni sustav (štitnjača, nadbubrežne žlijezde , hipofiza i drugi) nazivaju se endokrini. Trebalo bi detaljnije razmotriti žlijezde i njihove hormone.
Smješten ispred grkljana na vratu osobe, on proizvodi hormone bogate jodom - tiroksin, trijodotironin i kalcitonin. Ovi hormoni su dizajnirani za regulaciju metabolizma ugljikohidrata, masti i proteina u tijelu, a osim toga utječu na rast osobe i njen mentalni razvoj. Hiperfunkcijom ove žlijezde osoba razvija pretilost, tjelesna temperatura raste, a metabolizam se smanjuje.
To su male tvorbe iznad bubrega, koje se sastoje od dva vanjska (kortikalna) i unutarnja (cerebralna) sloja. Vanjski sloj proizvodi spolne hormone koji su odgovorni za razvoj spolnih karakteristika i reguliraju metabolizam (proteini i masti, ugljeni hidrati, kalijum, natrij). Disfunkcijom kore nadbubrežne žlijezde čovjek se razboli, takozvanom bronzanom bolešću, odnosno tijelo mu poprima bronzanu boju, a i sam doživljava stalni umor, gubitak apetita i mučninu. Funkcija moždine nadbubrežne žlijezde uključuje proizvodnju hormona poput noradrenalina i adrenalina. Dakle, adrenalin povećava brzinu otkucaja srca i ubrzava cirkulaciju krvi, mobilišući sve snage tijela u sebi stresna situacija... Ovog hormona ne nedostaje, ali njegov višak prijeti suženjem krvnih žila i ubrzanim otkucajima srca. Suprotno tome, norepinefrin je dizajniran da usporava rad srca.
Gušterača smještena u trbušnoj šupljini žlijezda je mješovite sekrecije, jer ima izvodne kanale i luči sekrete u želudac, sudjelujući u probavi. Jedan od hormona koji proizvodi, glikogen sintetiše glukozu, povećavajući tako nivo šećera u krvi. Drugi hormon, insulin, naprotiv, jedini smanjuje razinu šećera u krvi. Upravo zbog disfunkcije gušterače dijabetes, kod kojih dolazi do povećanja šećera u krvi.
Spolne žlijezde, odnosno ženski jajnici i muški testisi, također su žlijezde mješovite sekrecije. Zbog svog vanjskog lučenja pojavljuju se zametne stanice, odnosno spermatozoidi i jajašca, a njihova endokrina funkcija je stvaranje muških kao i ženskih spolnih hormona. Ovi hormoni utječu na seksualno ponašanje i lik osobe određenog spola. Muški testisi proizvode androgene koji su odgovorni za prisustvo muških spolnih karakteristika, mišića, brkova, brade, a ženski jajnici proizvode estrogene koji su odgovorni za spolne karakteristike lijepog spola, odnosno razvoj mliječnih žlijezda i oblik tijela.
Hipofiza koja se nalazi u mozgu sastoji se od 3 režnja (prednji, srednji i stražnji), od kojih dva proizvode hormone. U prednjem režnju stvara se hormon koji utječe na rast kostiju i ubrzava metaboličke procese. Njegov nedostatak može ugroziti razvoj patuljaštva, a hiperfunkcija može dovesti do gigantizma. Srednja hipofiza proizvodi hormon odgovoran za pigmentaciju kože.
Sve navedeno samo potvrđuje da su žlijezde i njihovi hormoni izuzetno važni za ljudsko tijelo i bez njih osoba ne bi mogla postojati. Trči sam!
Popularno na web mjestu
Informacije na našoj web stranici su informativne i edukativne prirode. Međutim, ove informacije ni na koji način nisu zamišljene kao vodič za samoliječenje. Obavezno se posavjetujte sa svojim liječnikom.
Da bismo razumjeli kako funkcioniraju endokrini organi, a posebno štitnjača, potrebno je ukratko razmotriti mehanizam djelovanja hormona.
Slika: 1. Izgled endokrinih organa
Endokrinu funkciju tijela pružaju sistemi koji uključuju:
1) endokrine žlezde koje luče hormone;
2) hormoni i različiti načini njihovog transporta;
3) relevantni organi ili ciljana tkiva koja reaguju na delovanje hormona.
Endokrini sistem održava postojanost unutrašnjeg okruženja tijela, što je neophodno za normalan tok fizioloških procesa.
Endokrine žlijezde su specijalizirani organi sa žljezdanom strukturom. Postoje žlijezde samo s unutrašnjim lučenjem (hipofiza, nadbubrežne žlijezde, štitnjača, paratireoidne žlijezde) i miješane - s unutarnjim i vanjskim lučenjem. Primjer je gušterača. Njegova vanjska sekrecija sastoji se u stvaranju probavnih enzima, koji posebnim kanalom ulaze u duodenum, a unutarnja sekrecija u proizvodnji hormona inzulina u specijaliziranim beta ćelijama otočića gušterače (Langerhans), koji ide direktno u krv i regulira šećer u krvi. ... Polne žlijezde također provode unutarnje i vanjske sekrecije.
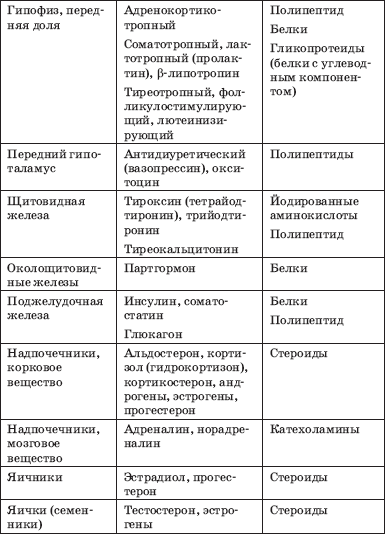
Naziv i položaj endokrinih žlijezda, hormoni koje oni proizvode i njihova kemijska priroda predstavljeni su u tablici. 1.
Tabela 1. Hormoni endokrinih žlijezda (Potemkin V.V., 1986)

Kraj tabele. 1
Izraz "hormon", preveden s grčkog, što znači "pobuditi", "inducirati", uveli su u praksu Bayliss i Starling. U januaru 1902. proveli su svoj poznati klasični eksperiment koji je uvjerljivo dokazao učešće humoralnog faktora u regulaciji sekretorne aktivnosti gušterače. Bayliss i Starling smatrali su da je hormon bilo koja supstanca koju normalno proizvode stanice svih dijelova tijela i prenosi je krv u udaljene dijelove, na kojima djeluje za dobro tijela u cjelini.
Trenutno su hormoni definirani kao visoko aktivne supstance koje nastaju u žlijezdama sa unutrašnjim lučenjem, ulaze u krvotok i vrše regulatorni efekat na funkcije organa i tjelesnih sistema udaljenih od mjesta njihovog lučenja. Oni se nazivaju i hemijskim glasnicima, koje specijalizovane ćelije izlučuju direktno u krvotok, koje su sposobne da sintetišu i oslobađaju hormone kao odgovor na određene signale.
Po hemijskoj strukturi hormoni se dijele na:
1) hormoni - derivati \u200b\u200baminokiselina;
2) proteinski i polipeptidni hormoni;
3) steroidni hormoni.
Prema svom fiziološkom učinku, hormoni se dijele na okidače i izvođače. Početni hormoni (aktivatori aktivnosti drugih endokrinih žlijezda) uključuju neurohormone hipotalamusa i tropske hormone hipofize. Izvršni hormoni imaju direktan utjecaj na osnovne funkcije tijela.
Hormoni se od ostalih biološki aktivnih supstanci razlikuju po sljedećim svojstvima:
1) vrlo visoka biološka aktivnost;
2) udaljena priroda radnje;
3) stroga specifičnost.
Visoku biološku aktivnost hormona karakterizira činjenica da, budući da su u krvi u zanemarljivim količinama, imaju izražen efekat.
Udaljena priroda djelovanja hormona leži u činjenici da su točke primjene njihovog djelovanja obično smještene daleko od mjesta stvaranja hormona u endokrinoj žlijezdi.
Hormoni se odlikuju strogom specifičnošću djelovanja. To znači da su reakcije organa, tkiva i ćelija na hormone strogo selektivne. Svaki hormon djeluje samo u određenim organima i tkivima, takozvanim ciljnim organima (ciljnim tkivima). Hormon prepoznaje i reagira sa svojim ciljnim organom jer u tim organima postoje posebna jedinjenja - receptori. Receptori su informativni molekuli proteina koji prepoznaju i transformiraju hormonalni signal u hormonsko djelovanje. Do danas je identificirano više od 60 receptora. Za steroide (hormone nadbubrežne kore) i hormone štitnjače (hormoni štitne žlijezde), koji lako prodiru kroz membranu, receptorski proteini se nalaze unutar ćelije. Receptori za proteinske hormone i kateholamine koji ne mogu proći kroz ćelijsku membranu nalaze se na površini ćelije.
Hipotalamus i hipofiza jedinstveni su sistem kontrole perifernih endokrinih žlijezda.
Hipotalamus je dio mozga koji ima svojstva nervnog i endokrinog sistema. Hipotalamus prima širok protok informacija od osjetila i unutrašnjih organa. Struktura neurosekretornih jezgara hipotalamusa uključuje takozvana velikoćelijska i maloćelijska jezgra. Prvi izlučuju hormone oksitocin i vazopresin koji se transportiraju duž nervnih trupaca do zadnjeg režnja hipofize, tamo se akumuliraju i prema potrebi koriste za regulaciju aktivnosti bubrega i maternice.
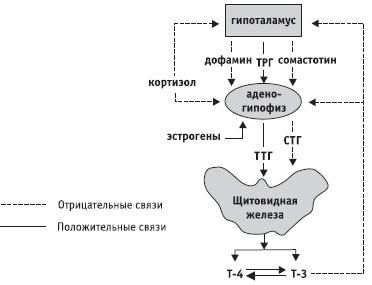
Slika: 2. Shema regulacije sistema hipotalamus-hipofiza-štitnjača
Ostale funkcije obavljaju jezgra malih ćelija hipotalamusa. Sposobni su da proizvode takozvane oslobađajuće hormone ili, tačnije, oslobađajuće faktore (rešavajući faktori). Faktori oslobađanja kroz venski sistem dopiru do hipofize i regulišu oslobađanje hormona potonje.
Regulacija aktivnosti hipofize hormonima maloćelijskih jezgara hipotalamusa vrši se prema antagonističkom principu. Jedna grupa faktora stimulira oslobađanje hipofiznih hormona (oslobađajući faktori ili liberini), a druga inhibira (statini). Poznati su sljedeći faktori: kortikoliberin, koji stimulira lučenje adrenokortikotropnog hormona hipofize; tiroliberin, koji pojačava oslobađanje hormona koji stimulira štitnjaču iz hipofize; somatoliberin i somatostatin (prvi stimulira oslobađanje somatotropnog hormona hipofize - hormona rasta, a drugi inhibira); melanoliberin i melanocitostatin, itd.
Hipofiza je središnja endokrina žlijezda u kojoj se proizvode takozvani tropski hormoni koji reguliraju funkciju perifernih žlijezda. To je složeni endokrini organ smješten u osnovi mozga - u takozvanom turskom sedlu. Sastoji se od adenohipofize, od kojih je većina prednji režanj žlijezde, i neurohipofize, predstavljene njezinim stražnjim režnjevom.
U prednjem režnju (adenohipofiza) proizvode se tropski hormoni:
Hormon rasta koji regulira rast tijela, sintezu proteina, razgradnju glukoze i masti;
Kortikotropin, koji stimulira sintezu glukokortikoida u nadbubrežnoj kori;
Tirotropin - stimulator sinteze tiroidnih hormona štitnjače;
Gonadotropin, folikulotropin, koji regulira sintezu muških i ženskih spolnih hormona;
Prolaktin je hormon koji regulira laktaciju.
U stražnjem režnju hipofize (neurohipofiza) nakupljaju se vazopresin i oksitocin. Vazopresin ili antidiuretski hormon regulira razmjenu vode i vaskularni tonus. Oksitocin povećava tonus glatkih mišića maternice, reguliše rad i lučenje mlijeka iz mliječnih žlijezda.
Periferne endokrine žlijezde podijeljene su u dvije skupine.
Prva se sastoji od žlijezda čija je funkcija regulirana tropskim hormonima adenohipofize. Zovu se žlijezde ovisne o adenohipofizi ili ciljane žlijezde. To uključuje štitnjaču, koru nadbubrežne žlijezde i endokrine dijelove spolnih žlijezda. Odnos adenohipofize i ciljnih žlijezda zasnovan je na principu "povratne sprege". Na primjer, adenohipofiza u krv izlučuje hormon koji stimulira štitnjaču, što stimulira lučenje hormona štitnjače tiroksina. Tiroksin koji je ušao u krv inhibira oslobađanje hormona koji stimulira štitnjaču iz hipofize.
Drugu grupu perifernih endokrinih žlijezda čine žlijezde čija funkcija ne ovisi o aktivnosti hipofize. Te se žlijezde nazivaju adenohipofizom ovisne. Oni funkcioniraju van mreže. Tu spadaju paratireoidne žlijezde, endokrini dio gušterače, medula nadbubrežne žlijezde i endokrine ćelije timusne žlijezde.
Timus (timus, ili timus, žlijezda) proizvodi hormone timozine i timopoetine - stimulanse imunoloških procesa.
Štitnjača proizvodi hormone koji sadrže jod: tiroksin i trijodotironin, kao i tirokalcitonin. Tiroksin i trijodtironin regulišu bazalni metabolizam, odnosno nivo potrošnje energije neophodan za održavanje vitalne aktivnosti tijela u stanju potpunog odmora. Tirokalcitonin reguliše razmjenu kalcijuma i fosfora.
U paratireoidnim žlijezdama proizvodi se paratiroidni hormon koji također regulira metabolizam kalcijuma i fosfora. Ali ako štitnjača tirokalcitonin smanji sadržaj kalcijuma u krvi, tada ga paratiroidni hormon povećava. Antagonistički odnos između tirokalcitonina i paratiroidnog hormona osigurava nivo kalcijuma u krvi na nivou neophodnom za tijelo.
Uloga hormona nadbubrežne žlijezde je izuzetno važna. To su upareni organi koji se nalaze iznad gornjih polova bubrega. U nadbubrežnim žlijezdama razlikuju se kora i moždina.
Kortikalna supstanca luči skupinu steroidnih hormona koji se zajednički nazivaju kortikosteroidi. Tri zone korteksa specijalizirane su za lučenje različitih hormona. Ćelije glomerularne zone proizvode mineralokortikoide: deoksikortikosteron i aldosteron, koji regulišu metabolizam minerala. Zona snopa proizvodi glukokortikoide: kortizol i kortikosteron, koji regulišu metabolizam proteina, masti i ugljenih hidrata. U retikularnom području sintetiziraju se neki prekursori muških spolnih hormona (androgeni).
Medula nadbubrežne žlijezde oslobađa kateholamine - adrenalin i norepinefrin - u krvotok. Norepinefrin djeluje ne samo kao hormon, već i kao posrednik u nervnim procesima simpatična podjela nervni sistem... Kateholamini imaju izražen vazokonstriktorni učinak, čime povećavaju krvni pritisak. Uključeni su u regulaciju metabolizma ugljikohidrata i masti, igraju glavnu ulogu u adaptaciji tijela za vrijeme stresa. Adrenalin se oslobađa kao odgovor na širok spektar podražaja: strah, uzbuđenje, bol, radost. Slikovito se naziva hitnim hormonom, hormonom emocija, prvim posrednikom stresa.
Insulin, glukagon, somatostatin proizvode se u endokrinoj gušterači (Langerhansovi otočići). Insulin je najvažniji regulator ugljenih hidrata, kao i metabolizma masti i proteina. Glukagon je fiziološki antagonist insulina, kao i stimulator sekrecije insulina u prisustvu glukoze. Somatostatin inhibira lučenje insulina, glukagona i hormona rasta. Kršenje sekrecije insulina i glukagona dovodi do razvoja tako ozbiljne i raširene bolesti kao što je dijabetes melitus.
Polne žlijezde proizvode ne samo hormone, već i spolne ćelije (spermu i jajašca). Testisi (testisi) proizvode muške spolne hormone - androgene, od kojih je glavni testosteron. Androgeni doprinose razvoju primarnih i sekundarnih muških spolnih karakteristika. U jajnicima se sintetišu ženski spolni hormoni - estrogeni koji su odgovorni za stvaranje ženskih primarnih i sekundarnih spolnih karakteristika, kao i progesteron, hormon neophodan za normalan tok trudnoće. Proizvodnja hormona i zametnih ćelija vrši se pod kontrolom gonadotropnih hormona adenohipofize.
Bubrezi, koji vrše funkciju izlučivanja, takođe su vrsta endokrinih žlijezda. Ćelije takozvanog jukstaglomerularnog aparata bubrega izlučuju u krv hormon renin koji je uključen u stvaranje angiotenzina II, najaktivnijeg regulatora vaskularnog tonusa. Bubrezi takođe proizvode eritropoetin, hormon koji stimuliše stvaranje crvenih krvnih zrnaca u koštanoj srži.
Utvrđeno je da je srce takođe endokrina žlijezda. U pretkomori se sintetizira natriuretski hormon koji utječe na izlučivanje natrijuma bubrezima.
Endokrini organ koji privremeno funkcionira je posteljica ("bebino mjesto"). Proizvodi hormone koji doprinose normalnom toku trudnoće.
U centralnom nervnom sistemu stvaraju se posebne supstance - neuroendokrini peptidi (neurohormoni) - endorfini, enkefalini. Nazivaju se "endogeni opijati" ili morfiju slični peptidi. Ovi hormoni imaju analgetičke (ublažavajuće bolove) efekte i reprodukuju efekte morfija na ponašanje.
Jedinstvo i međusobna povezanost nervnog i endokrinog mehanizma vrlo se jasno prati na primjeru funkcioniranja hipotalamo-hipofiznog sistema. Trenutno je ispravnije govoriti ne o endokrinom sistemu, već o neuroendokrinom sistemu tijela.
Iznijevši opće ideje o endokrinim žlijezdama, prijeđimo na glavni cilj naše priče - štitnjaču.
Pitanje broj 2 Endokrine žlijezdei njihovi hormoni.
Endokrine žlijezde(od grčkog endon - iznutra, crio - izdvajam) ili endokrine žlijezde, su specijalizirani organi ili skupine ćelija, čija je glavna funkcija razviti i osloboditi određene biološki aktivne tvari u unutarnju okolinu tijela. Endokrine žlijezde nemaju izvodne kanale. Njihove ćelije su opletene bogatom mrežom krvnih i limfnih žila, a otpadni proizvodi se ispuštaju direktno u krv, limfu i tkivnu tečnost. Ova karakteristika u osnovi razlikuje endokrine žlijezde od egzokrinih žlijezda koje luče svoje izlučevine kroz kanale za izlučivanje.
Proizvodi koje proizvode žlijezde sa unutrašnjim lučenjem nazivaju se hormoni(Grčki hormao - uzbuditi, aktivirati). Izraz "unutrašnja sekrecija" predložio je 1885. godine francuski fiziolog C. Bernard, a izraz "hormon" - engleski fiziolozi W. Beilis i E. Starling 1902. godine.
Sljedeće osobine su karakteristične za hormone: Njihovu sintezu i izolaciju provode specijalizirane ćelije. Hormoni se formiraju u žljezdanim endokrinim ćelijama, nakon čega ulaze u unutrašnje okruženje, uglavnom u krv i limfu. Intermedijarni proizvodi sinteze ili metabolizma hormona često imaju biološku aktivnost, ali u pravilu se ne selektiraju .
Visoka biološka aktivnost.Hormoni imaju fiziološki učinak u vrlo niskim koncentracijama. Dakle, koncentracija ženskog spolnog hormona (estradiola) u krvi kreće se od 0,2 do 0,6 μg (10 -6 g) u 100 ml plazme. Sadržaj hormona rasta u krvi mjeri se u još manjim količinama - nanogramima (KG 9 g). Hipofiza reagira na pikograme (10 -12 g) hormona hipotalamusa, angiotenzin-P-proizvod endokrinih bubrežnih ćelija - u femtogramima (10 -15 g) izaziva osjećaj žeđi. Osim hormona, nijedan drugi hemijski otpad nije efikasan u tako malim dozama.
Specifičnost.Svaki hormon karakterizira određena, njemu samo svojstvena hemijska struktura, mjesto sinteze i funkcija. S tim u vezi, nedostatak bilo kojeg hormona ne može se nadoknaditi drugim hormonima ili biološki aktivnim supstancama.
Akcija na daljinu.Hormoni se, po pravilu, prenose krvlju daleko od mjesta nastanka, utječući na udaljene organe i tkiva. Po tome se razlikuju od medijatora i citokina koji djeluju na jednu ćeliju ili skupinu ćelija na mjestu njihovog stvaranja.
Hemijska struktura hormona i njihovitransformacija u tijelu
Prema svojoj hemijskoj strukturi, hormoni se mogu podijeliti u 4 glavne skupine: proteini i peptidi, derivati \u200b\u200baminokiselina, steroidi, prostaglandini.
Primjeri proteinskih hormona: inzulin, somatotropin (hormon rasta), tropski hormoni prednje hipofize. Neki od njih (folitropin, tirotropin, lutropin) su složeni proteini, drugi (insulin, kalcitonin, itd.) Su jednostavni proteini. Struktura peptida ima glukagon, vazopresin, oksitocin, hipotalamičke hormone. Derivati \u200b\u200baminokiselina uključuju hormone štitnjače - trijodotironin, tiroksin, kao i adrenalin i norepinefrin. Steroidni hormoni temelje se na cikličnoj ugljikovodičnoj jezgri ciklopentaneperhidrofenantrena. Ova grupa uključuje hormone nadbubrežne kore i spolne žlijezde.
Glavne faze formiranja i transformacije hormona mogu se predstaviti na sljedeći način:
biosinteza hormona;
lučenje, tj. izolacija iz endokrine ćelije;
transport krvi u periferna tkiva;
prepoznavanje hormonskog signala od strane ciljnih ćelija;
transdukcija (prevođenje) hormonskog signala u biološki odgovor;
suzbijanje hormonskog signala.
Organi i tkiva sa endokrinom funkcijom ćelije
|
|
|
|
Organi i tkiva
|
|
|
|
|
|
Placenta
|
Sincitiotrofoblasti Citotrofoblasti
|
Horionski gonadotropin, prolaktin Estriol, progesteron
|
|
|
|
|
Timociti
|
Timozin, timopoetin
|
|
|
|
|
Intersticijum Cjevčice
|
Eritropoetin Kalcitriol
|
|
|
|
|
Atrijalni miociti
|
Atriopeptid, somatostatin, angiotenzin-P
|
|
|
|
|
|
|
|
Organi, tkiva i ćelije sa endokrinom funkcijom
|
|
|
|
Strukture
|
Tkiva, ćelije
|
Hormoni
|
|
|
|
Hipotalamus
|
Živčane ćelije struktura malih točaka
|
Hipotalamični neurohormoni:
|
|
|
|
|
Stražnji hipotalamus Živčane ćelije velikih ćelijskih jezgara prednjeg hipotalamusa
|
a) liberini: kortikoliberin, tiroliberin, luliberin, folliberin, somatoliberin, prolaktoliberin, melanoliberin; b) statini: prolaktostatin, melanostatin, somatostatin Vazopresin, oksitocin
|
|
|
|
Adenohipofiza Neurohipophysis
|
Kortikotrofi Gonadotrofi Thyrotrophs Somatotrofi Lactotrophs Pituicites
|
Kortikotropin, melanotropin Folitropin, lutropin Tirotropin Hormon rasta Prolaktin Vazopresin, oksitocin
|
|
|
|
Nadbubrežne žlijezde Cortex Mozak je važan
|
Zona glomerula Zona snopa Mrežasta zona Kromafinske stanice
|
Mineralokortikoidi Glukokortikoidi Seksualni steroidi: androgeni, estrogeni Andrenalin, noradrenalin
|
|
|
|
Štitnjača
|
Folikularni tirociti
|
Trijodotironin, tetrajodotironin Kalcitonin, katakalcin, Ko-kalcigenin
|
|
|
|
Paratireoidne žlijezde
|
Glavne ćelije K-ćelija
|
Paratirin kalcitonin
|
|
|
|
|
Pineociti
|
Melatonin
|
|
|
|
Gušterača
|
Langerhansovi otoci: ćelije ose R-ćelije A-ćelije
|
Glukagon Somatostatin
|
|
|
|
Spolne žlijezde (testisi, jajnici)
|
Leydigove ćelije Sertolijeve ćelije Granulozne ćelije Corpus luteum
|
Testosteron estrogeni, inhibin estradiol, estron, progesteron Progesteron
|
|
|
|
|
|
|
|
Biosinteza hormonaprogramiran u genetskom aparatu specijalizovanih endokrinih ćelija. Stoga to ovisi o strukturi i ekspresiji gena koji kodiraju sintezu ovih hormona, kao i o enzimima koji reguliraju sintezu hormona i post-translacijske procese. Odsustvo ili defekt odgovarajućih gena dovodi do endokrinopatije. Primjer je patuljak s genetskim nedostatkom u hormonu rasta.
2. Lučenje hormona.najvažnija karakteristika svakog hormona je njegovo lučenje. Pakovan u vezikule ili granule, hormon se prenosi prema citoplazmatskoj membrani. Otpuštanju proteinsko-peptidnih hormona i kateholamina iz ćelije prethodi interakcija citoplazmatske membrane i membrane sekretorne granule. Nakon toga dolazi do njihove lize i hormon se oslobađa iz ćelije. Ovaj proces aktiviraju mnogi faktori, posrednici, visoka koncentracija kalijuma, električni podražaji itd. Izlučivanje hormona čin je praćen trošenjem energije, stoga je uvijek povezan sa pomacima u ATP-cAMP sistemu. Za sekreciju je neophodno učešće kalcijumovih jona koji aktiviraju proteine \u200b\u200bmikrotubularno-mikrofilamentnog sistema, olakšavajući interakciju hormonskih granula s njima i utječu na stvaranje cAMP. Stoga smanjenje sadržaja jona kalcijuma u izvanstaničnoj okolini i njegov ulazak u endokrinu ćeliju neizbežno dovodi do smanjenja sekretorne aktivnosti ove ćelije.
3.
Transport hormona.Izlučeni hormon ulazi u unutrašnje okruženje tijela, uglavnom u krv, i dalje se prenosi. Većina hormona stvara složene spojeve s proteinima plazme u krvi. Neki od ovih proteina su specifični transportni proteini (na primjer, transkortin, koji veže hormone nadbubrežne kore), a neki su nespecifični (na primjer, y-globulini). Kompleksiranje s proteinima je reverzibilan proces. Uz to, neki od hormona u krvi povezani su s tijelima, posebno eritrociti.
Stvaranje vezanog oblika hormona od velike je fiziološke važnosti. Prvo, štiti tijelo od viška nakupljanja u krvi (i, prema tome, utjecaja na tkiva) slobodnih hormona. Drugo, pridruženi oblik hormona je njegova fiziološka rezerva. Treće, vezivanje proteina pomaže u zaštiti hormona od razgradnje enzimima, tj. produžava mu život. Konačno, kompleksacija s proteinima sprečava filtraciju hormona male molekulske težine kroz bubrežne glomerule i na taj način ograničava ove važne regulatorne procese.
4.
Prepoznavanje hormonskog signala.Nakon ulaska u periferne organe, hormoni se, u pravilu, oslobađaju iz proteinske komponente i, fiksirajući se na određene receptore ćelija koje percipiraju dati hormon (ciljne ćelije), provode svoje specifično djelovanje. U procesu perifernog djelovanja hormona dolazi do njihovih različitih transformacija. U ovom je slučaju moguće stvaranje novih hormonalnih proizvoda, često aktivnijih ili koji proizvode drugačiji biološki efekt od izvornog hormona. Dakle, tiroidni hormon tiroksin može se u tkivima pretvoriti u trijodotironin, aktivniji hormon iste žlijezde. Androgeni (muški spolni hormoni) pretvaraju se u hipotalamusu u estrogene - ženske spolne hormone. Transdukcija hormonskog signala u biološki odgovor organsko je povezana s mehanizmom djelovanja ovog hormona.
Djelovanje bilo kojeg hormona na ciljne ćelije uvijek započinje njegovom interakcijom s određenim komponentama ćelije. Ova pojava naziva se prijem hormona, a stanične komponente koje djeluju s hormonom nazivaju se receptori. Receptori hormona su kiseli oligopeptidi velike molekulske težine.
Strukturu molekula receptora karakterizira asimetrija. Postoje tri odjeljka:
hormon vezanja;
efektor, prenoseći hormonski signal na unutarćelijske mehanizme, tj. pretvaranje signala u biološki odgovor;
povezivanje prvog i drugog odjeljka.
- hipofiza sintetiše / luči hormon rasta (STH), prolaktin, ACTH, itd .;
- u nadbubrežnoj žlijezdi postoje četiri sloja ćelija, od kojih svaka sintetizira svoj vlastiti hormon.
S gledišta gastroenterologa, gušterača je egzokrini organ, jer luči enzime gušterače; sa stanovišta endokrinologa, on je endokrini organ, jer proizvodi paket međuovisnih hormona (insulin, glukagon, somatostatin, itd.).
Pored toga, neki hormoni se proizvode na nekoliko mjesta:
- kateholamini - ne samo u medulli nadbubrežne žlijezde, već i u ganglijima paravertebralnog živca;
- somatostatin - i na Langerhansovim otočićima i u hipotalamusu.
Izvan endokrinih žlijezda pronađene su mikroskopske nakupine ćelija koje su se specijalizirale za sintezu biološki aktivnih supstanci sa hormonskim svojstvima:
- regulatori lučenja hormona endokrinih žlijezda:
- jezgra hipotalamusa sintetišu supstance koje regulišu lučenje hormona hipofize (somatoliberin, hormon koji oslobađa ACTH, itd.);
- nakupine ćelija u crijevnom zidu koje proizvode inkretinske hormone;
- regulatori funkcija organa:
Relativno nedavno otkrivene su biološki aktivne supstance leptin i adiponektin, sintetizovane masnim tkivom (adipociti), koje su klasifikovane kao hormoni, s obzirom da imaju sistemsko regulatorno dejstvo - regulišu apetit i metabolizam energije.
Dakle, hormone proizvodi ne samo endokrina žlijezda, uslijed čega ova kvaliteta ne može jednoznačno definirati pojam "hormona". Istodobno, u modernoj kliničkoj endokrinologiji gotovo sve bolesti predstavljaju jedno ili drugo kršenje funkcija endokrine žlijezde. S tim u vezi, definicija hormona i s tim povezana definicija endokrine žlijezde u kliničkoj endokrinologiji i dalje je "klasična".
Dakle, možemo dati slijedeću, s kliničke točke gledišta sasvim cjelovitu definiciju hormona.
Hormon - biološki aktivna supstanca koju proizvodi endokrina žlijezda, a koja ima regulatorni učinak na određene strukture tijela i metabolizam (iskorištavanje supstrata iz krvi, razmjena energije itd.), što se često očituje vanjskim vidljivim promjenama u tijelu (na primjer, rastom) i / ili promjenama u ponašanju na primjer, seksualno).
U ovoj klasičnoj definiciji pojmovi endokrina žlijezda i hormon međusobno su ovisni. Stoga je logika dijagnostičkog pretraživanja u kliničkoj endokrinologiji očigledna - kroz proučavanje hormona krvi za dijagnosticiranje bolesti endokrinih žlijezda.
Definicija endokrine žlijezde
Endokrina žlijezda - dobro definirana makroanatomska struktura čija je glavna funkcija sinteza biološki aktivnih supstanci zvanih hormoni. U kliničkoj endokrinologiji razlikuje se sedam endokrinih žlijezda čije se funkcije procjenjuju ispitivanjem hormona koje žlijezda proizvodi u krvi. Za procjenu njegovih funkcija koristi se ne čitav spektar hormona žlijezda, već strogo ograničen skup njih, uz pomoć kojih se određuje funkcija endokrine žlijezde. Pored hormona, njihovi metaboliti mogu se koristiti za dijagnozu bolesti, koje se ponekad pokažu pouzdanijim markerom endokrinih bolesti od proučavanja samih hormona. Dakle, u dijagnozi feokromocitoma, proučavanje metanefrinskih metabolita kateholamina pouzdanije je od adrenalina i norepinefrina.
Proučavanje hormona za dijagnozu endokrinih bolesti nije uvijek opravdano. Najupečatljiviji je primjer dijabetes melitus, kod čije se dijagnoze ne koristi testiranje insulina, iako je bolest uzrokovana nedostatkom insulina. Takođe, studija oksitocina i vazopresina ne koristi se za dijagnozu njihovog nedovoljnog ili prekomjernog lučenja, a kršenje njihove sinteze određeno je njihovim metaboličkim učincima.
Štoviše, u dijagnozi endokrinih bolesti mogu se koristiti hormoni koje endokrine žlijezde ne sintetiziraju, na primjer, faktor rasta sličan insulinu (IGF-I), koji se stvara u jetri pod djelovanjem STH. Koristi se za dijagnozu akromegalije uzrokovane tumorom hipofize.
Sinteza hormona od strane endokrine žlijezde može biti:
- njegova jedina funkcija (na primjer, prednji režanj hipofize);
- u kombinaciji sa stvaranjem zametnih ćelija (na primjer, jajnika i testisa);
- u kombinaciji s egzokrinom sekrecijom (na primjer, gušterača);
- u kombinaciji sa taloženjem hormona koji se sintetišu izvan njega.
Endokrina žlijezda može sintetizirati:
- jedini hormon koji je rijedak (na primjer, paratiroidna žlijezda);
- spektar hormona (obično):
- specijaliziranim ćelijskim podstrukturama, posebno u nadbubrežnim žlijezdama, dvije ćelijske podstrukture - kora i medula - proizvode steroidne hormone, odnosno kateholamine;
- odvojene ćelije, ujedinjene ili ne u izoliranim kompleksima, na primjer, u hipofizi, određeni hormoni sintetiziraju pojedinačne ćelije koje nisu kombinirane u različite ćelijske formacije; u gušterači insulin i glukagon proizvode β- i α-ćelije, ujedinjene u Langerhansovim otočićima.
Priroda i funkcija hormona
Hormoni su podijeljeni u dvije glavne skupine.
Polipeptidi ili derivati \u200b\u200baminokiselina (većina):
- složeni polipeptidi (LH, hCG);
- peptidi srednje veličine;
- mali peptidi;
- dipeptidi (T4 i T3);
- derivati \u200b\u200bpojedinih aminokiselina (serotonin, histamin).
Derivati \u200b\u200bholesterola su steroidi dvije vrste:
- s netaknutim steroidnim prstenom (steroidi nadbubrežnih žlijezda i spolnih žlijezda);
- s nepovezanim prstenom B.
Postoje četiri glavne funkcije hormona u tijelu:
- reprodukcija;
- rast i razvoj;
- proizvodnja, korišćenje i očuvanje energije.
Odvojeni hormon, s jedne strane, može imati različite biološke efekte na različite organe, au jednom organu i na različito vrijeme; s druge strane, neki su biološki procesi pod integralnom kontrolom nekoliko hormona.
Hormoni reguliraju funkcije svojih sljedećih ciljeva:
- druge endokrine žlijezde (npr. hipofiza-nadbubrežna žlijezda);
- funkcionalni sistemi;
- organi (npr. T4 i funkcija srca ili T4 i funkcija mozga);
- tkiva (kao što su kortizol i kosti).
Sinteza, skladištenje i lučenje hormona
Peptidni hormoni se sintetišu istim mehanizmom kao i bilo koji drugi proteini. Često se prvo sintetizira velika molekula prohormona, koja se zatim pretvara u manji hormon. Na primjer, pretparatireoidni hormon → proparatiroidni hormon → paratireoidni hormon. S druge strane, steroidi i kateholamini se sintetišu iz manjih molekula.
Endokrini organi nisu jedinstveno mjesto za sintezu hormona, međutim, samo se u njima sinteza hormona i njihova regulacija odvijaju najefikasnije. Tri glavne karakteristike razlikuju endokrini organ od ne-endokrinog tkiva koje sintetiše bilo koji hormon:
- brzina sinteze je mnogo veća u endokrinom organu;
- endokrine žlijezde su opremljene mehanizmom za transport hormona u krv, koji je obično reguliran.
Brzina izlučivanja hormona žlijezdom određuje se brzinom njegove sinteze, koju mogu regulirati drugi hormoni tropski u ovu žlijezdu. Izuzev T4 i 1,25-dihidroksiholekalciferola, zalihe hormona u tijelu su vrlo ograničene.
Poticanje lučenja hormona povezano je s depolarizacijom ćelijske membrane i otvaranjem kalcijumovih kanala, što dovodi do ulaska kalcijuma u ćeliju, gdje se kombinira s proteinom koji veže kalcij.
Transport i eliminacija hormona
Hormoni se uklanjaju iz krvi kao rezultat metaboličkih procesa, na primjer, peptidni hormoni se inaktiviraju proteolitičkim enzimima. U jetri se hormoni kombiniraju s glukuronskom kiselinom i luče se žučom, ali se istovremeno djelomično reapsorbiraju i uključuju se u takozvani enterohepatični ciklus. Hormoni se takođe izlučuju urinom.
Mali molekuli hormona (posebno T 4) vežu se za proteine \u200b\u200bu krvi, što usporava njihovo izlučivanje iz krvi i održava malu bazu slobodnih hormona u krvi na potrebnom nivou. Vezivanje na proteine \u200b\u200btakođe olakšava transport steroida topivih u mastima.
Hormonski receptori
Hormonski receptori su ćelijski proteini koji vezuju hormon.
Interakcija s hormonom uzrokuje konformacijsku promjenu receptora, što aktivira određeni sistem ćelijskih enzima, koji zapravo ostvaruje karakteristični učinak hormona. Kada se hormon veže za receptor ćelijske membrane, u citozolu se pojavljuju takozvani drugi glasnici (prvi je hormon). U ćelijskoj jezgri kompleks hormona i receptora vezan je za deoksiribonukleinsku kiselinu (DNK) i reguliše ekspresiju gena. Maksimalni učinak hormona obično se pojavljuje čak i ako je vezano manje od 50% receptora. Slobodni receptori oslobođeni veze sa hormonom ponovo se vraćaju u citosol ili u ćelijsku membranu, gdje nastavljaju sudjelovati u interakciji hormona i receptora.
Steroidni hormoni su lipofilni, stoga se slobodno difundiraju kroz ćelijsku membranu, a zatim se vežu za proteine \u200b\u200bcitosolnih receptora.
T3 se veže za proteine \u200b\u200bnuklearnih receptora, a kompleks T3-receptora, povezujući se sa DNK, stimulira stvaranje glasničke RNK. Steroidni i tiroidni hormoni često djeluju sinergijski, međusobno pojačavajući specifične efekte (pojačavajući ekspresiju gena).
Broj receptora ćelijske membrane i unutarćelijskih receptora se mijenja, a mijenja se i snaga njihove veze sa hormonom. Ćelije miometrija i mlečnih žlezda sadrže receptore za oksitocin, čiji se broj povećava pod dejstvom estrogena (regulacija prema gore), a smanjuje se pod dejstvom progesterona (regulacija dole). U miokardu se nalaze norepinefrinski receptori (β 1), čiji se broj i afinitet za norepinefrin povećavaju pod dejstvom hormona štitnjače (T 3 / T 4).
Vodotopivi hormoni (monoamini, aminokiseline i peptidi) vezuju se za receptore membrane koja je zasićena lipidima, pa ne dopušta da hormoni topivi u vodi slobodno difundiraju kroz membranu. U hormonalnoj reakciji ćelije, vodotopivi hormoni nazivaju se prvim glasnicima. Kao odgovor na njihovu interakciju s receptorom unutar ćelije, aktiviraju se takozvani drugi glasnici - cAMP, ciklični gvanozin monofosfat, inositol trifosfat, joni kalcijuma, diacilglicerol, itd. Kalcijumovi ioni služe kao vrlo važan drugi glasnik. Protok jona kalcijuma kroz ćelijsku membranu u citosol kontrolira se komunikacijom hormona i receptora, nervnim podražajima ili ga modificiraju drugi drugi glasnici.
Koncentracija hormona u većini slučajeva je 10 -10 mol / l. U ovom slučaju, vezivanje jedne molekule za membranski receptor dovodi do stvaranja 10 000 cAMP molekula u ćeliji, i u tom pogledu cAMP djeluje kao molekularno pojačalo hormonskog signala (10 000 puta!). Fosfodiesteraza uništava cAMP, pa njegovi inhibitori teofilin i kofein djeluju sinergijski s hormonima, u kojima je cAMP drugi glasnik. cAMP stimulira kataboličke procese - lipolizu, glikogenolizu (glukagon), glukoneogenezu i ketogenezu, lučenje insulina u β-ćelijama i pankreasu.
Sažetak na temu
Endokrine žlijezde i njihovi hormoni
Endokrine žlijezde
(od grčkog endon - iznutra, crio - izdvajam) ili endokrine žlijezde, su specijalizirani organi ili skupine ćelija, čija je glavna funkcija razvijanje i oslobađanje specifičnih biološki aktivnih tvari u unutarnju okolinu tijela. Endokrine žlijezde nemaju izvodne kanale. Njihove ćelije su opletene obilnom mrežom krvnih i limfnih žila, a otpadni proizvodi se ispuštaju direktno u krv, limfu i tkivnu tečnost. Ova karakteristika u osnovi razlikuje endokrine žlijezde od egzokrinih žlijezda, koje luče svoje izlučevine kroz izvodne kanale. Proizvodi koje proizvode žlijezde sa unutrašnjim lučenjem nazivaju se hormoni(Grčki hormao - uzbuditi, aktivirati). Izraz "unutrašnja sekrecija" predložio je 1885. godine francuski fiziolog C. Bernard, a izraz "hormon" - engleski fiziolozi W. Beilis i E. Starling 1902. godine.
Sljedeće osobine su karakteristične za hormone: Njihovu sintezu i izolaciju provode specijalizirane ćelije.
Hormoni se formiraju u žlezdanim endokrinim ćelijama, nakon čega ulaze u unutrašnje okruženje, uglavnom u krv i limfu. Intermedijarni proizvodi sinteze ili metabolizma hormona često imaju biološku aktivnost, ali u pravilu se ne selektiraju .
Visoka biološka aktivnost.
Hormoni imaju fiziološki učinak u vrlo niskim koncentracijama. Dakle, koncentracija ženskog spolnog hormona (estradiola) u krvi kreće se od 0,2 do 0,6 μg (10 -6 g) u 100 ml plazme. Sadržaj hormona rasta u krvi mjeri se u još manjim količinama - nanogramima (KG 9 g). Hipofiza reagira na pikograme (10 -12 g) hormona hipotalamusa, angiotenzin-P-proizvod endokrinih bubrežnih ćelija - u femtogramima (10 -15 g) izaziva osjećaj žeđi. Osim hormona, niti jedan drugi hemijski otpad nije efikasan u tako malim dozama. Specifičnost.Svaki hormon karakterizira specifična, svojstvena samo hemijska struktura, mjesto sinteze i funkcija. S tim u vezi, nedostatak bilo kojeg hormona ne može se nadoknaditi drugim hormonima ili biološki aktivnim supstancama. Akcija na daljinu.
Hormoni se, po pravilu, prenose krvlju daleko od mjesta nastanka, utječući na udaljene organe i tkiva. Po tome se razlikuju od medijatora i citokina koji djeluju na jednu ćeliju ili skupinu ćelija na mjestu njihovog stvaranja.
Hemijska struktura hormona i njihova transformacija u tijelu
Prema svojoj hemijskoj strukturi, hormoni se mogu podijeliti u 4 glavne skupine:
proteini i peptidi;
derivati \u200b\u200baminokiselina;
steroidi;
prostaglandini.
Primjeri proteinskih hormona uključuju inzulin, somatotropin (hormon rasta), tropske hormone prednje hipofize. Neki od njih (folitropin, tirotropin, lutropin) su složeni proteini, drugi (insulin, kalcitonin, itd.) Su jednostavni proteini. Struktura peptida ima glukagon, vazopresin, oksitocin, hipotalamičke hormone. Derivati \u200b\u200baminokiselina uključuju hormone štitnjače - trijodotironin, tiroksin, kao i adrenalin i norepinefrin. Steroidni hormoni temelje se na cikličnoj ugljikovodičnoj jezgri ciklopentaneperhidrofenantrena. Ova grupa uključuje hormone nadbubrežne kore i spolne žlijezde. Glavne faze formiranja i transformacije hormona mogu se predstaviti na sljedeći način:
biosinteza hormona;
lučenje, tj. izolacija iz endokrine ćelije;
transport krvi u periferna tkiva;
prepoznavanje hormonskog signala od strane ciljnih ćelija;
transdukcija (prevođenje) hormonskog signala u biološki odgovor;
suzbijanje hormonskog signala.
Organi i tkiva sa ćelijskom endokrinom funkcijom
№
|
Organi i tkiva
|
Ćelije
|
Hormoni
|
1.
|
Placenta
|
Sincitiotrofoblasti Citotrofoblasti
|
Horionski gonadotropin, prolaktin Estriol, progesteron
|
2.
|
Timus
|
Timociti
|
Timozin, timopoetin
|
3.
|
Bud
|
YUGA Intersticijum
Cjevčice
|
Renin Eritropoetin
Kalcitriol
|
4.
|
Srce
|
Atrijalni miociti
|
Atriopeptid, somatostatin, angiotenzin-P
|
Organi, tkiva i ćelije sa endokrinom funkcijom
№
|
Strukture
|
Tkiva, ćelije
|
Hormoni
|
1.
|
Hipotalamus
|
Živčane ćelije struktura malih točaka
|
Hipotalamični neurohormoni:
|
|
|
|
Stražnji hipotalamus Živčane ćelije velikih staničnih jezgara prednjeg hipotalamusa
|
a) liberini: kortikoliberin, tiroliberin, luliberin, folliberin, somatoliberin, prolaktoliberin, melanoliberin; b) statini: prolaktostatin, melanostatin, somatostatin
Vazopresin, oksitocin
|
2.
|
Hipofiza Adenohipofiza Neurohipophysis
|
Kortikotrofi Gonadotrofi
Thyrotrophs
Somatotrofi
Lactotrophs
Pituicites
|
Kortikotropin, melanotropin Folitropin, lutropin Tirotropin Hormon rasta Prolaktin
Vazopresin, oksitocin
|
3.
|
Nadbubrežne žlijezde Kortikalna supstanca
Mozak je važan
|
Zona glomerula Zona snopa Mrežasta zona Kromafinske stanice
|
Mineralokortikoidi Glukokortikoidi Seksualni steroidi: androgeni, estrogeni Andrenalin, noradrenalin
|
4.
|
Štitnjača
|
Folikularni tirociti K ćelije
|
Trijodotironin, tetrajodotironin Kalcitonin, katakalcin, Ko-kalcigenin
|
5.
|
Paratireoidne žlijezde
|
Glavne ćelije K-ćelija
|
Paratirin kalcitonin
|
6.
|
Epifiza
|
Pineociti
|
Melatonin
|
7.
|
Gušterača
|
Langerhansovi otoci: ćelije ose R-ćelije A-ćelije
|
Glukagon Insulin
Somatostatin
|
8.
|
Spolne žlijezde (testisi, jajnici)
|
Leydigove ćelije Sertolijeve ćelije Granulozne ćelije Corpus luteum
|
Testosteron estrogeni, inhibin estradiol, estron, progesteron Progesteron
|
Biosinteza hormonaprogramiran u genetskom aparatu specijalizovanih endokrinih ćelija. Posljedično, to ovisi o strukturi i ekspresiji gena koji kodiraju sintezu ovih hormona, kao i o enzimima koji reguliraju sintezu hormona i post-translacijske procese. Odsustvo ili nedostatak odgovarajućih gena dovodi do endokrinopatije. Primjer je patuljak s genetskim nedostatkom u hormonu rasta.
2. Lučenje hormona.Kao što je naznačeno, najvažnija karakteristika svakog hormona je njegovo lučenje. Pakiran u vezikule ili granule, hormon se prenosi prema citoplazmatskoj membrani. Otpuštanju proteinsko-peptidnih hormona i kateholamina iz ćelije prethodi interakcija citoplazmatske membrane i membrane sekretorne granule. Nakon toga dolazi do njihove lize i hormon se oslobađa iz ćelije. Ovaj proces aktiviraju mnogi faktori, posrednici, visoka koncentracija kalijuma, električni podražaji itd. Izlučivanje hormona čin je praćen trošenjem energije, stoga je uvijek povezan sa pomacima u ATP-cAMP sistemu. Za sekreciju je neophodno učešće kalcijumovih jona koji aktiviraju proteine \u200b\u200bmikrotubularno-mikrofilamentnog sistema, olakšavajući interakciju hormonskih granula s njima i utječu na stvaranje cAMP. Stoga smanjenje sadržaja jona kalcijuma u izvanstaničnoj okolini i njegov ulazak u endokrinu ćeliju neizbežno dovodi do smanjenja sekretorne aktivnosti ove ćelije. 3.
Transport hormona.Izlučeni hormon ulazi u unutrašnje okruženje tijela, uglavnom u krv, i dalje se prenosi. Većina hormona stvara složene spojeve s proteinima plazme u krvi. Neki od ovih proteina su specifični transportni proteini (na primjer, transkortin, koji veže hormone nadbubrežne kore), a neki su nespecifični (na primjer, γ-globulini). Kompleksiranje s proteinima je reverzibilan proces. Uz to, neki od hormona u krvi povezani su s tijelima, posebno eritrociti. Stvaranje vezanog oblika hormona od velike je fiziološke važnosti. Prvo, štiti tijelo od viška nakupljanja u krvi (i, prema tome, utjecaja na tkiva) slobodnih hormona. Drugo, pridruženi oblik hormona je njegova fiziološka rezerva. Treće, vezivanje proteina pomaže u zaštiti hormona od razgradnje enzimima, tj. produžava mu život. Konačno, kompleksacija s proteinima sprečava filtraciju hormona male molekulske težine kroz bubrežne glomerule i na taj način ograničava ove važne regulatorne procese.
4.
Prepoznavanje hormonskog signala.Nakon ulaska u periferne organe, hormoni se, u pravilu, oslobađaju iz proteinske komponente i, fiksirajući se na određene receptore ćelija koje percipiraju dati hormon (ciljne ćelije), provode svoje specifično djelovanje. U procesu perifernog djelovanja hormona dolazi do njihovih različitih transformacija. U ovom je slučaju moguće stvaranje novih hormonalnih proizvoda, često aktivnijih ili koji proizvode drugačiji biološki efekt od izvornog hormona. Dakle, tiroidni hormon tiroksin može se u tkivima pretvoriti u trijodotironin, aktivniji hormon iste žlijezde. Androgeni (muški spolni hormoni) pretvaraju se u hipotalamusu u estrogene - ženske spolne hormone. Transdukcija hormonskog signala u biološki odgovor organsko je povezana s mehanizmom djelovanja ovog hormona. Djelovanje bilo kojeg hormona na ciljne ćelije uvijek započinje njegovom interakcijom s određenim komponentama ćelije. Ova pojava naziva se prijem hormona, a stanične komponente koje djeluju s hormonom nazivaju se receptori. Receptori hormona su kiseli oligopeptidi velike molekulske težine.
Strukturu molekula receptora karakterizira asimetrija. Postoje tri odjeljka:
hormon vezanja;
efektor, prenoseći hormonski signal na unutarćelijske mehanizme, tj. pretvaranje signala u biološki odgovor;
povezivanje prvog i drugog odjeljka.
Receptori se nalaze ili unutarćelijski ili na površini citoplazmatske membrane. Unutarćelijski receptori vezuju steroidne i tiroidne hormone, membranski receptori vezuju ostale hormone. Također se vjeruje da funkciju prepoznavanja specifičnog hormonskog signala u svim stanicama za sve hormone vrši membranski receptor, a nakon što se hormon veže za odgovarajući receptor, daljnja uloga hormonsko-receptorskog kompleksa za peptidne i steroidne hormone je drugačija. U peptidu, proteinskim hormonima i kateholaminima, hormon-receptorski kompleks dovodi do aktivacije membranskih enzima i stvaranja različitih sekundarnih glasnika (glasnika) hormonskog regulatornog efekta, koji svoje djelovanje ostvaruju u citoplazmi, organelama i ćelijskom jezgru.
Poznata su četiri sistema sekundarnih posrednika:
adenilat ciklaza - ciklični adenozin monofosfat (cAMP);
gvanilat ciklaza - ciklični gvanozin monofosfat (cGMP);
fosfolipaza C - inozitol trifosfat (ITP);
jonizovani kalcijum.
4.1.
Sistem "adenilat ciklaza - cAMP".
Membrane enzim adenilat ciklaza mogu biti u dva oblika - aktivirani i neaktivirani. Aktivacija adenilat ciklaze događa se pod utjecajem hormonsko-receptorskog kompleksa, čije stvaranje dovodi do vezivanja gvanil nukleotida (GTP) s posebnim regulatorno stimulirajućim proteinom (GS-protein), nakon čega GS-protein uzrokuje vezivanje magnezijuma za adenilat-ciklazu i njegovu aktivaciju. Na taj način hormoni koji aktiviraju adenilat ciklazu - glukagon, tirotropin, paratirin, vazopresin, gonadotropin itd. Neki hormoni, naprotiv, suzbijaju adenilat ciklazu (somatostatin, angiotenzin-P, itd.). Pod uticajem adenilat ciklaze, cAMP se sintetiše iz ATP, koji aktivira protein kinaze u citoplazmi ćelije, koje pružaju fosforilaciju brojnih unutarćelijskih proteina. To mijenja propusnost membrane, tj. uzrokuje metaboličke i, shodno tome, funkcionalne promjene tipične za hormon. Unutarćelijski efekti cAMP-a takođe se manifestuju u uticaju na procese proliferacije, diferencijacije, na dostupnost proteina membranskih receptora molekulima hormona.
Gvanilat ciklaza - cGMP sistem.
Aktivacija membranske gvanilat-ciklaze ne događa se pod direktnim uticajem hormonsko-receptorskog kompleksa, već indirektno putem jonizovanog sistema kalcijuma i oksidansa. Tako atrijalni natriuretski hormon - atriopeptid, tkivni hormon vaskularnog zida - ostvaruje svoje učinke. U većini tkiva biohemijski i fiziološki efekti cAMP i cGMP su suprotni. Primjeri uključuju stimulaciju srčanih kontrakcija pod utjecajem cAMP i inhibiciju njihovog cGMP, stimulaciju kontrakcija glatkih mišića crijeva cGMP i suzbijanje cAMP.
4.3 Sistem "fosfolipaza C - jonositol trifosfat".
Kompleks hormona i receptora uz učešće regulatornog proteina G dovodi do aktivacije membranskog enzima fosfolipaze C, što uzrokuje hidrolizu membranskih fosfolipida formirajući dva sekundarna prenosnika - inositol fosfat i diacilglicerol. Iiositol trifosfat uzrokuje oslobađanje jonizovanog kalcijuma iz unutarćelijskih zaliha. Potonji, vežući se za protein kalmodulin, aktivira protein kinaze i fosforilaciju unutarćelijskih strukturnih proteina i enzima. Dijacilglicerol takođe aktivira i dovršava proces fosforilacije drugih proteina, istovremeno ostvarujući drugi put hormonskog efekta: stvaranjem arahidonske kiseline, koja je izvor supstanci snažnih u smislu metaboličkih i fizioloških efekata - prostaglandina i leukotriena. Kroz razmatrane sisteme sekundarnih medijatora ostvaruju se efekti adrenalina, vazopresina, angiotenzina-P, somatostatina, oksitocina i nekih drugih hormona.
4.4. Sistem "kalcijum - kalmodulin".
Jonizovani kalcijum ulazi u ćeliju nakon stvaranja hormonsko-receptorskog kompleksa ili iz vanstaničnog okruženja uslijed aktiviranja sporih kalcijumovih kanala (na primjer u miokardu), ili iz unutarćelijskih zaliha pod utjecajem gore opisanih unutarćelijskih procesa. U citoplazmi nemišićnih ćelija kalcijum se veže za poseban protein - kalmodulin, a u mišićnim ćelijama ulogu kalmodulina ima troponin C. Kalmodulin vezan za kalcijum aktivira brojne protein kinaze koje pružaju fosforilaciju proteina. Kratkoročni porast količine kalcijuma u ćeliji i njegovo vezivanje s kalmodulinom poticaj je za brojne fiziološke procese - kontrakciju mišića, lučenje i oslobađanje medijatora hormona, sintezu DNK, promjene u mobilnosti ćelija, aktivnost enzima i transport supstanci kroz membrane. U steroidnim hormonima, membranski receptor pruža specifično prepoznavanje hormona i njegov transfer u ćeliju. U citoplazmi postoji poseban receptor za citoplazmatski protein s kojim se hormon veže. Ova veza s receptorskim proteinom neophodna je za ulazak steroidnog hormona u jezgru, gdje on stupa u interakciju s trećim nuklearnim receptorima, vezivanjem kompleksa "hormon - nuklearni receptor" sa akceptorima hromatina, specifičnim kiselim proteinom i DNK, što podrazumijeva aktivaciju transkripcije mRNA, sintezu transporta i ribosomska RNA, transport mRNA u citoplazmu, translacija mRNA uz sintezu proteina i enzima u ribosomima. Svi ovi fenomeni zahtijevaju dugotrajno prisustvo hormonsko-receptorskog kompleksa u jezgri. Međutim, učinci steroidnih hormona pojavljuju se ne samo nakon nekoliko sati, neki se brzo, u roku od nekoliko minuta. To su efekti poput povećanja membranske propusnosti, povećanog transporta glukoze i aminokiselina, oslobađanja lizosomskih enzima, pomaka u energiji mitohondrija i, uz to, povećanja cAMP i jonizovanog kalcijuma. Dakle, stav je potpuno opravdan, čiji pristaše vjeruju da membranski receptor za steroidne hormone ne samo da vrši funkciju „prepoznavanja“ molekula hormona, već poput receptora za peptidni hormon aktivira sistem sekundarnih glasnika u ćeliji. Peptidni hormoni takođe imaju sposobnost selektivnog uticaja na transkripciju gena u ćelijskom jezgru. Ovaj se učinak može ostvariti ne samo sa ćelijske površine zbog sekundarnih medijatora, već i prilivom hormona u ćeliju uslijed internetizacijakompleks hormona i receptora (zbog endocitoze).
Fenomen internalizacije hormonsko-receptorskih kompleksa, a time i smanjenje broja hormonskih receptora na staničnoj membrani omogućava nam da shvatimo mehanizam smanjenja osjetljivosti efektora na prekomjernu količinu hormonskih molekula, ili fenomen desenzibilizacijaefektor. Ova pojava je u stvari negativna regulatorna povratna informacija na nivou efektora. Suprotan fenomen je senzibilizacija,ili povećanje osjetljivosti na hormone, može biti posljedica povećanja broja slobodnih mjesta receptora na membrani, kako zbog smanjenja internalizacije, tako i kao rezultat "plutanja" aktivnih mjesta vezivanja receptora. Dakle, hormoni prenose informativne signale ćeliji, a sama ćelija je sposobna regulirati stepen percepcije hormonske kontrole. 5. Transdukcija hormonskog signala u biološki odgovor.Postoji 5 vrsta hormonskog djelovanja na ciljno tkivo: metaboličko, morfogenetsko, kinetičko, korektivno, reaktogeno. Metaboličko djelovanje.
Metaboličke promjene uzrokovane hormonima temelje se na promjenama u funkciji ćelija, tkiva i organa. Morfogenetsko dejstvo -
uticaj hormona na procese formiranja, diferencijacije i rasta strukturnih elemenata. Primjeri su utjecaj hormona rasta na rast tijela i unutrašnjih organa, utjecaj spolnih hormona na razvoj sekundarnih spolnih karakteristika. Kinetička akcija
- sposobnost hormona da pokreću aktivnost efektora, da uključuju provođenje određene funkcije. Na primjer, oksitocin uzrokuje kontrakciju mišića maternice, adrenalin pokreće razgradnju glikogena u jetri i oslobađanje glukoze u krv, vazopresin aktivira mehanizam reapsorpcije vode u sabirnim kanalima nefrona. Korektivna radnja
- promjene u aktivnosti organa ili procesi koji se javljaju u odsustvu hormona. Primjeri korektivnog djelovanja hormona su utjecaj adrenalina na brzinu otkucaja srca, aktiviranje oksidativnih procesa tiroksinom i smanjenje reabsorpcije kalijumovih jona u bubrezima pod utjecajem aldosterona. Svojevrsno korektivno djelovanje je normalizirajući učinak hormona, kada je njihov utjecaj usmjeren na obnavljanje izmijenjenog ili čak poremećenog procesa. Na primjer, s početnom prevalencijom anaboličkih procesa metabolizma proteina, glukokortikoidi uzrokuju katabolički učinak, ali ako u početku prevladava razgradnja proteina, glukokortikoidi stimuliraju njihovu sintezu. Reaktogeno dejstvo
- sposobnost hormona da promijeni reaktivnost tkiva na djelovanje istog hormona, drugih hormona ili posrednika nervnih impulsa. Na primjer, hormoni štitnjače pojačavaju efekte kateholamina, hormoni koji regulišu kalcijum smanjuju osjetljivost distalnog nefrona na djelovanje vazopresina. Varijacija reaktogenog djelovanja hormona je permisivno djelovanje, koje podrazumijeva sposobnost jednog hormona da omogući ostvarenje učinka drugog hormona. Tako, na primjer, glukokortikoidi djeluju permisivno u odnosu na kateholamine (za ostvarenje efekata adrenalina neophodno je prisustvo malih količina kortizola); insulin ima permisivan efekat na somatotropin, itd. 6. Suzbijanje hormonskog signala.Ovaj mehanizam djeluje na normalizaciju hormonskog signala (ako se odgodi, postojat će opasnost od prekomjerne napetosti u mnogim funkcijama). Brojni su mehanizmi za suzbijanje hormonskog signala. Prije svega, ovo je smanjenje biosintetske i sekretorne aktivnosti samih ćelija koje proizvode hormone. Zajedno s tim događa se i uništavanje (katabolizam) nekih izlučenih hormona. Kao rezultat, značajan udio hormona nestaje iz krvi nakon što se oslobode iz granula. Enzimi jetre inaktiviraju hormone deaminacijom i metilacijom. U procesu inaktivacije, hormoni i proizvodi njihovog katabolizma mogu se kaugirati u jetri glukuronskom i sumpornom kiselinom, što povećava njihovu topljivost u vodi i olakšava naknadno izlučivanje kroz bubrege. Neki od slobodnih i konjugovanih hormona ulaze u žuč i izlučuju se iz tijela kroz crijeva. Suzbijanje hormonskog signala olakšava se desenzibilizacijom gore opisanih tkiva sa dugotrajnim povišenim nivoom hormona u krvi. U nekim slučajevima nastaju antitijela na hormone i drugi inhibitori koji inhibiraju biološki efekat.
Smanjenje aktivnosti hormonskog signala postiže se i kao rezultat lučenja kontraregulacijskih hormona, tj. hormoni suprotnog djelovanja. Na primjer, višak insulina uzrokuje hipoglikemiju, koja zauzvrat inducira lučenje kontrainzularnih hormona. Drugi važan fiziološki mehanizam za gašenje hormonskog signala je inhibicija sekrecije prema principu povratne sprege.
Savremena teorija regulatornih peptida
Regulatorni peptidi- biološki aktivne supstance koje sintetišu ćelije tela različitog porekla i učestvuju u regulaciji različitih funkcija. Među njima se razlikuju neuropeptidi koje izlučuju nervne ćelije i koji su uključeni u provođenje funkcija nervnog sistema. Pored toga, nalaze se izvan centralnog nervnog sistema u brojnim endokrinim žlijezdama, kao i u drugim organima i tkivima.
U ontogeniji su se regulatorni peptidi pojavili mnogo ranije od „klasičnih“ hormona, tj. prije izolacije specijaliziranih endokrinih žlijezda. To nam omogućava da pretpostavimo da je odvojeno stvaranje navedenih skupina supstanci programirano u genomu, te su stoga neovisne.
Izvori regulatornih peptida su pojedinačne ćelije koje proizvode hormon, a ponekad čine male nakupine. Te ćelije se smatraju početnim oblikom endokrinih formacija. Tu spadaju neurosekretorne ćelije hipotalamusa, neuroendokrine (hromafinske) ćelije nadbubrežnih žlijezda i paraganglija, stanice sluznice gastrointestinalnog sistema, pinealociti epifize. Utvrđeno je da su ove ćelije sposobne za dekarboksilaciju aromatičnih kiselina-prekursora neuroamina, što je omogućilo njihovo kombiniranje u jedinstveni sistem (Pearse, 1976), nazvan „APUD-sistem“ (od prvih slova engleskih riječi Unos prekursora amina i sistem dekarboksilacije - prethodnici amina). Veliki broj peptida (vazoaktivni intestinalni peptid - VIP, holecistokinin, gastrin, glukagon) u početku je pronađen u sekretornim elementima gastrointestinalnog trakta. Ostali (supstanca P, neurotenzin, enkefalini, somatostatin) izvorno su pronađeni u nervnom tkivu. Treba napomenuti da su u gastrointestinalnom traktu neki peptidi (gastrin, holecistokinin, VIP i neki drugi) prisutni u živcima, kao i u endokrinim ćelijama.
Postojanje ovog neurodifuznog endokrinog sistema objašnjava se migracijom ćelija iz jednog izvora - neuronskog grebena; uključeni su u središnji živčani sistem i tkiva različitih organa, gdje se transformiraju u stanice slične središnjem živčanom sistemu, lučeći neuroamine (neurotransmitere) i peptidne hormone. To objašnjava prisustvo neuropeptida u crijevima i gušterači, ćelija Kulčitskog u bronhima, a također jasno objašnjava pojavu hormonski aktivnih tumora pluća, crijeva i gušterače. Apudociti se nalaze i u bubrezima, srcu, limfnim čvorovima, koštanoj srži, epifizi i posteljici.
Glavne skupine regulatornih peptida (prema Kriegeru)
Najčešća klasifikacija regulatornih peptida uključuje sljedeće grupe:
hormoni koji oslobađaju hipotalamus;
hormoni neurohipofize;
hipofizni peptidi (ACTH, MSH, STH, TSH, prolaktin, LH, FSH, (3-endorfin, lipotropini);
gastrointestinalni peptidi;
ostali peptidi (angiotenzin, kalcitonin, neuropeptid V).
Za određeni broj peptida utvrđena je lokalizacija stanica koje sadrže i distribucija vlakana. Opisano je nekoliko peptidergičnih moždanih sistema koji su podijeljeni u dva glavna tipa.
Sistemi za dugu projekciju,
čija vlakna dopiru do udaljenih dijelova mozga. Na primjer, tijela neurona iz porodice proopiomelanokortina nalaze se u lučnom jezgru hipotalamusa, a njihova vlakna dopiru do amigdale i periakveduktalne sive materije srednjeg mozga.
Sustavi kratkih projekcija:
tijela neurona su često smještena u mnogim dijelovima mozga i imaju lokalnu raspodjelu procesa (supstanca P, enkefalini, holecistokinin, somatostatin).
Mnogi su peptidi prisutni u perifernim živcima. Na primjer, supstanca P, VIP, enkefalini, holecistokinin, somatostatin nalaze se u vagusnim, celijakijskim i išijasnim živcima. Medula nadbubrežne žlijezde sadrži velike količine preproenkefalina A (metenkefalina). Pokazano je postojanje neuropeptida i neurotransmitera u istom neuronu: serotonin se nalazi u neuronima produljene moždine zajedno sa supstancom P, dopaminom, zajedno s holecistokininom, u neuronima srednjeg mozga, acetilholinom i VIP-om, u autonomnim ganglijima. O funkcionalnom značaju ovog suživota mogu se suditi prema sljedećim faktorima. Pod utjecajem VIP-a u fiziološkim koncentracijama dolazi do izraženog povećanja osjetljivosti muskarinskih receptora na acetilholin u submandibularnoj žlijezdi mačaka, a antiserum VIP-u djelomično blokira vazodilataciju izazvanu stimulacijom parasimpatičkih živaca.
Sinteza regulatornih peptida.
Karakteristična karakteristika sinteze peptida je njihovo stvaranje fragmentacijom velike molekule preteče, tj. kao rezultat takozvanog post-translacijskog proteolitičkog cijepanja - obrade. Sinteza prekursora događa se u ribosomima, što potvrđuje prisustvo glasničke RNK koja kodira peptid i post-translacijske modifikacije enzima s oslobađanjem aktivnih peptida - u Golgijevom aparatu. Ovi peptidi do aksonskog transporta stižu do nervnih završetaka.
Aktivni peptidi koji potiču iz jednog prekursora čine njegovu porodicu. Opisane su sledeće porodice peptida.
Porodica proopiomelanokortina (POMC).Neuronska tijela u kojima je prisutan ovaj veliki protein (286 aminokiselinskih ostataka) lokalizirana su u lučnom jezgru hipotalamusa. Ovisno o skupu enzima iz POMK-a formiraju se: u prednjem režnju hipofize - uglavnom ACTH, (3-lipotropin, P-endorfin, u međuproduktu - c-melanostimulirajući hormon i P-endorfin. Dakle, skup enzima određuje specijalizaciju proizvodnje ćelijama strogo definiranih pe To su enzimi katepsin B, tripsin, karboksipeptidaza, aminopeptidaza, mjesta njihovog napada su upareni aminokiselinski ostaci.
Porodica Cerulein:gastrin, holecistokinin.
VIP porodica: secretin, glukagon.
Porodica arginin-vazopresin:vazopresin, oksitocin.
Pored toga, utvrđeno je da met-enkefalin i leu-enkefalin imaju prekursore u obliku preproenkefalina A, odnosno preproenkefalina B. Proteoliza u ovom slučaju nije inaktivacija, već transformacija aktivnosti. Mehanizam djelovanja neuropeptida:
Karakteristična karakteristika regulatornih peptida je polifunkcionalnost (u smislu mehanizma i prirode efekata) i formiranje regulatornih lanaca (kaskade). Generalno, mehanizmi djelovanja peptida mogu se podijeliti u dvije skupine: sinaptičke i ekstrasinaptičke. 1.
Sinaptički mehanizmi djelovanja peptidamože se izraziti u neurotransmiteru ili neuromodulatornoj funkciji. Neurotransmiter (Peyrotransmiter)- tvar koja se oslobađa od presinaptičkog terminala i djeluje na sljedeću - postsinaptičku membranu, tj. vrši prenosnu funkciju. Utvrđeno je da neki peptidi obavljaju ovu funkciju putem peptidergijskih receptora smještenih na neuronima (njihovim tijelima ili terminalima). Tako se luteinizirajući hormon hipotalamički oslobađajući hormon (luliberin) u sinaptičkim ganglijima žabe oslobađa nakon stimulacije živca kroz proces ovisan o kalcijumu i uzrokuje kasni polagani ekscitacijski postsinaptički potencijal.
Za razliku od "klasičnih" neurotransmitera (noradrenalin, dopamin, serotonin, acetilholin), peptidi koji obavljaju transfernu funkciju odlikuju se velikim afinitetom receptora (koji može pružiti udaljeniji efekat) i produženim (desetinama sekundi) delovanjem zbog odsustva sistema za inaktivaciju enzima i povratni depozit.
Neuromodulator,za razliku od neurotransmitera, on ne uzrokuje neovisni fiziološki efekat u postsinaptičkoj membrani, ali modificira reakciju ćelije na neurotransmiter. Dakle, neuromodulacija nije prijenosna funkcija, već regulatorna funkcija koja se može izvoditi i na post- i presinaptičkom nivou. Vrste neuromodulacije:
kontrola otpuštanja neurotransmitera sa terminala;
regulacija cirkulacije neurotransmitera;
modifikacija efekta "klasičnog" neurotransmitera.
2.
Ekstra-sinaptičko djelovanje peptidaimplementiran na nekoliko načina. A. Parakrina akcija(parakrinija) - provodi se u područjima međustaničnog kontakta. Na primjer, somatostatin, koji luče A-ćelije otočnog tkiva gušterače, obavlja parakrinu funkciju u kontroli lučenja insulina i glukagona (za 3- i α-ćelije, odnosno kalcitonin - kod kontrole sekrecije hormona koji sadrže jod štitnjača). B. Neuroendokrino djelovanje- sprovodi se oslobađanjem peptida u krvotok i njegovim efektom na efektorsku ćeliju. Primjeri uključuju somatostatin i druge hipotalamičke faktore, koji se luče u medijalnoj eminenciji s nekih terminala u portalnu cirkulaciju i kontroliraju lučenje hormona hipofize. B. Endokrino djelovanje.U ovom slučaju, peptidi se oslobađaju u opšti krvotok i djeluju kao udaljeni regulatori. Ovaj mehanizam uključuje komponente potrebne za "klasične" endokrine funkcije - transportne proteine \u200b\u200bi receptore ciljne ćelije. Utvrđeno je da se kao stabilizatori koriste: neurofizini - za vazopresin i oksitocin, neki albumini i globulini u plazmi - za holecistokinin i gastrin. Što se tiče recepcije, utvrđeno je postojanje zasebnih receptora za opioidne peptide, vazopresin, VIL. Ciklični nukleotidi, proizvodi hidrolize fosfoinozitida, kalcijuma i kalmodulina sa naknadnom aktivacijom protein kinaze i kontrolom fosforilacije translacionih i transkripcionih regulacijskih proteina mogu se koristiti kao sekundarni glasnici. Pored toga, opisan je mehanizam internalizacije, kada regulatorni peptid, zajedno s receptorom, uđe u ćeliju putem mehanizma sličnog pinocitozi, a signal se prenese na genom neurona. Regulatorni peptidi karakterizirani su stvaranjem složenih lanaca ili kaskada kao rezultat činjenice da su metaboliti nastali iz glavnog peptida također funkcionalno aktivni. Ovo objašnjava trajanje efekata kratkotrajnih peptida.
Funkcije regulatornih peptida: 1. Bol.Brojni peptidi utječu na stvaranje bola kao složenog psihofiziološkog stanja tijela, uključujući sam osjećaj bola, kao i emocionalne, voljne, motoričke i autonomne komponente. U ovom slučaju, peptidi su uključeni u nociceptivni i antinociceptivni sistem. Tako se supstanca P, somatostatin, VIP, holecistokinin i angiotenzin nalaze u primarnim senzornim neuronima, a supstanca P je neurotransmiter koji luče određene klase aferentnih neurona. Istovremeno, enkefalini, vazopresin, angiotenzin i srodni opioidni peptidi nalaze se u silaznom supraspinalnom putu koji vodi do stražnjih rogova kičmene moždine i vrše inhibitorni efekat na nociceptivne puteve (analgetički efekat).
2. Sjećanje, učenje, ponašanje.Bilo je dokaza da nedostaju fragmenti ACTH (ACTH 4-7 i ACTH 4-10) hormonalni efekti, i ss-melanostimulirajući hormon poboljšavaju kratkoročno pamćenje, a vazopresin je uključen u stvaranje dugoročnog pamćenja. Ubrizgavanje antitela na vazopresin u moždane komore u roku od sat vremena nakon treninga izaziva zaborav. Pored toga, ACTH 4-10 poboljšava pažnju.
Utvrđen je uticaj određenog broja peptida na ponašanje u ishrani. Primjeri uključuju pojačavanje motivacije za hranu pod djelovanjem opioidnih peptida i slabljenje pod djelovanjem holecistokinina, kalcitonina i kortikoliberina.
Opioidni peptidi imaju značajan učinak na emocionalne reakcije, budući da su endogeni euforigen.
VIP ima hipnotičko, hipotenzivno i bronhodilatacijsko dejstvo. Threoreoberin ima psihotonski efekat. Luliberin, osim što vrši zapovjednu funkciju (stimulacija gonadotropa prednje hipofize), regulira seksualno i roditeljsko ponašanje.
3. Vegetativne funkcije.Razni peptidi uključeni su u kontrolu nivoa krvni pritisak... Ovo je sistem renin-angiotenzin, čije su sve komponente prisutne u mozgu, opioidni peptidi, VIP, kalcitonin, atriopeptid, koji imaju snažno natriuretsko djelovanje.
Opisane su promjene u termoregulaciji pod djelovanjem nekih peptida. Dakle, intracentralna primjena tiroliberina i Rβ-endorfin uzrokuje hipertermiju, dok primjena ACTH i α-MSH indukuje hipotermiju.
4. Stres.Značajno je da su brojni neuropeptidi (opioidni peptidi, prolaktin, epifize peptida) klasificirani kao antistresni sistem, jer ograničavaju razvoj stresnih reakcija. Tako je u eksperimentima s različitim modelima pokazano da opioidni peptidi ograničavaju aktivaciju simpatikusa nervnog sistema i svih veza hipotalamičko-hipofizno-nadbubrežnog sistema, sprečavajući iscrpljivanje tih sistema, kao i neželjene posljedice viška glukokortikoida (suzbijanje upalne reakcije i timusno-limfni sistem, pojava čirevi gastrointestinalni trakt i drugi) - Antihipotalamični faktori epifize inhibiraju stvaranje liberina i lučenje hormona prednje hipofize. Smanjena aktivacija hipotalamusa ograničava hipersekreciju vazopresina, što štetno djeluje na miokardij.
5. Uticaj na imunološki sistem.Uspostavljene su dvosmjerne veze između regulatornog peptidnog sistema i imunološkog sistema. S jedne strane, sposobnost mnogih peptida da moduliraju imunološki odgovor dovoljno je proučena. Suzbijanje sinteze imunoglobulina pod dejstvom (3-endorfina, enkefalina, ACTH i kortizola; inhibicija sekrecije interleukina -1
(IL -1)
i razvoj groznice pod uticajem hormona stimulišućeg melanocite. Utvrđeno je da vazoaktivni crijevni peptid (VIL) inhibira sve funkcije limfocita i njihovo oslobađanje iz limfnih čvorova, što se smatra novim oblikom imunomodulacije. U isto vrijeme, brojni peptidi djeluju stimulirajuće na imunološki sistem, uzrokujući povećanje sinteze imunoglobulina i γ-interferona (| 3-endorfin, hormon koji stimulira štitnjaču), povećanje aktivnosti prirodnih ćelija ubica (R.-endorfin, enkefalini), povećanje proliferacije limfocita i oslobađanje limfokina (supstanca P, prolaktin, hormon rasta), povećanje proizvodnje superoksidnih aniona (hormon rasta). Opisani su receptori limfocita za brojne hormone. S druge strane, imunološki medijatori utječu na metabolizam i izlučivanje hipotalamičkih neurotransmitera i oslobađanje hormona. Dakle, regulatorni leukopeptid IL -1
sposoban je prodrijeti u mozak kroz područja povećane propusnosti krvno-moždane barijere i stimulirati lučenje hormona koji oslobađa kortikotropin (u prisustvu prostaglandina), nakon čega slijedi stimulacija oslobađanja ACTH i kortizola, koji inhibiraju stvaranje IL -1
i imunološki odgovor.
Istovremeno, oslobađanjem somatostatina, IL -1
inhibira lučenje TSH i hormona rasta. Dakle, imunopeptid djeluje kao okidač, koji zatvaranjem mehanizma povratne sprečavanja sprečava suvišnost imunološkog odgovora.
Prema modernim konceptima, kompletan regulatorni krug između neuroendokrinih i imunoloških mehanizama takođe uključuje peptide zajedničke za oba sistema. Konkretno, prikazana je sposobnost hipotalamičkih neurona da luče IL-1. Izoliran je gen odgovoran za njegovu proizvodnju, čiju ekspresiju indukuju bakterijski antigeni i kortikotropin. Opisani su neuronski putevi u mediobazalni hipotalamus čovjeka i pacova koji sadrži IL-1 i IL-6, kao i ćelije hipofize koje luče ove peptide.
Dakle, imunološki medijatori mogu regulirati funkcije prednjeg dijela hipofize putem:
endokrini mehanizam (limfokini aktiviranih limfocita koji cirkulišu u krvi);
neuroendokrini efekti koje realizuju interleukini hipotalamusa kroz tuberoinfundibularni portalni sistem;
parakrina kontrola u samoj hipofizi.
S druge strane, rezultati imunohemijskih i molekularnih studija pokazali su da imunokompetentne ćelije luče mnogo peptida i hormona povezanih sa endokrinom i neuronskom aktivnošću: limfociti i makrofagi sintetišu ACTH; limfociti - hormon rasta, prolaktin, TSH, enkefalini; mononuklearni limfociti i mastociti - VIP, somatostatin; ćelije timusa - arginin, vazopresin, oksitocin, neurofizin. U ovom slučaju, hormoni hipofize koje luče limfociti regulirani su istim faktorima kao i hipofiza. Na primjer, lučenje ACTH limfocitima inhibiraju glukokortikoidi i stimuliraju hormon koji oslobađa kortikotropin. Predložen je koncept prema kojem oslobađanje navedenih hormona od strane limfocita osigurava autokrinu i parakrinu regulaciju lokalnog imunološkog odgovora. Dakle, funkcije tri glavna regulatorna sistema - nervnog, endokrinog i imunološkog - integrirane su u složene regulatorne krugove koji funkcioniraju prema principu povratne sprege. Istovremeno, periferni limfociti, ako slijedimo koncept D. Blalocka (Blalock, 1989), pružaju senzorni mehanizam putem kojeg se prepoznaju nekognitivni podražaji (strane tvari) i mobiliziraju neuroendokrini adaptivni odgovori.
Učešće regulatornih peptida u razvoju patologije.
Budući da peptidni hormoni čine polifunkcionalni sistem koji je uključen u regulaciju mnogih funkcija u tijelu, vjerovatno je da će biti uključeni u patogenezu različitih bolesti. Dakle, kršenje koncentracije moždanih peptida utvrđeno je kod degenerativnih neuroloških bolesti nepoznate etiologije: Alzheimerove bolesti (smanjenje koncentracije somatostatina u moždanoj kori) i Huntingtonove bolesti (smanjenje koncentracije holecistokinina, supstance P i enkefalina, povećanje sadržaja somatostatina, bazalnih ganglija, kao bazalnih ganglija vezujući holecistokinin u tim strukturama i u kori velikog mozga). Hoće li ove promjene biti primarne ili se pojavljuju kao posljedica razvoja bolesti, ostaje za vidjeti.
Otkriće opioidnih peptida i distribucija njihovih receptora u različitim moždanim strukturama, posebno u limbičkom sistemu, skrenulo je pažnju na procenu njihovog značaja u patogenezi mentalnih poremećaja. Predlaže se hipoteza o postojanju nedostatka opioida u bolesnika sa shizofrenijom, posebno o nemogućnosti stvaranja γ-endorfina koji ima neuroleptički učinak. Utvrđeno je povećanje koncentracije atriopeptida tokom stagnacije u cirkulacijskom sistemu, što je, možda, mehanizam za kompenzaciju kršenja metabolizma natrijuma (njegovo kašnjenje).
Proučavanje oligopeptidnih hormona kao regulatornog sistema dovelo je do izolacije posebne grupe bolesti prouzrokovanih njegovom patologijom - apudopatije.
Apudopatija- bolesti povezane sa kršenjem strukture i funkcije apudocita i izražene u određenim kliničkim sindromima. Postoje primarne apudopatije uzrokovane patologijom samih apudocita, a sekundarne nastaju kao reakcija apudocita na kršenje homeostaze u tijelu uzrokovane bolešću čija patogeneza nije prvenstveno povezana s patologijom APUD sistema (kod zaraznih bolesti, rasta tumora, bolesti nervnog sistema itd.) ). Primarne apudopatije mogu se manifestirati hiperfunkcijom, hipofunkcijom, disfunkcijom i stvaranjem apudoma - tumora iz ćelija APUD sistema. Primjeri su sljedeći apudomi.
Gastrinoma- apudom iz ćelija koje proizvode gastrin, za koji je poznato da potiče lučenje veliki broj želučani sok visoke kiselosti i probavne snage. Stoga se klinički gastrinoma očituje razvojem ulcerogenih Zollinger Ellison sindroma. Kortikotropinoma- apudoma, koji se razvija iz apudoblasta gastrointestinalnog trakta i manifestuje se ektopičnom hiperprodukcijom ACTH i razvojem Itsenko-Cushingovog sindroma. Vipoma- tumor iz ćelija koji luče vazoaktivni crevni peptid. Lokalizirano u dvanaesniku ili gušterači. Manifestira se razvojem vodene dijareje i dehidracije, kao i poremećajem metabolizma elektrolita. Somatostatinoma- tumor iz crijevnih ćelija ili otočnog tkiva gušterače, proizvodeći somatostatin. Somatostatinoma se obično razvija kao tumor D ćelija gušterače koje luče somatostatin. Karakterizira ga klinički sindrom koji uključuje dijabetes melitus, žučna bolest, hipohlorhidrija, steatoreja i anemija. Dijagnosticira se povećanjem koncentracije somatostatina u krvnoj plazmi. Primena regulatornih peptida u medicini
Na osnovu regulatornih peptida, neki lijekovi... Dakle, oligopeptidi (kratki peptidi) N-terminalnog fragmenta ACTH i MSH koriste se za ispravljanje pažnje i pamćenja, vazopresin - za poboljšanje pamćenja kod traumatičnih i drugih amnezija. Domaći lijek dalargin (analog lejenkefalina) široko se koristi u medicinskoj praksi. Započela je komercijalna proizvodnja surfagona (analoga luliberina), namijenjenog korekciji poremećaja reproduktivnog sistema. |